




Cruzar la papa que se cultiva con fines comerciales con algunas de las cientos de especies silvestres puede mejorar la resistencia de la planta a factores adversos así como las propiedades culinarias de sus tubérculos.
Desde hace años los investigadores están tratando de mejorar las cualidadesde la papa cultivada cruzándola con algunas de sus cientos de especies silvestres emparentadas, muchas de las cuales han desarrollado resistencia a agentes externos adversos y propiedades culinarias que pueden transferirse a las papas de uso comercial.
La papa que se cultiva comercialmente -Solanum tuberosum subespecie tuberosum- está emparentada con más de doscientas especies silvestres que, como ella, poseen tallos subterráneos suculentos (denominados tubérculos), ricos en almidón, proteinas y vitamina C.
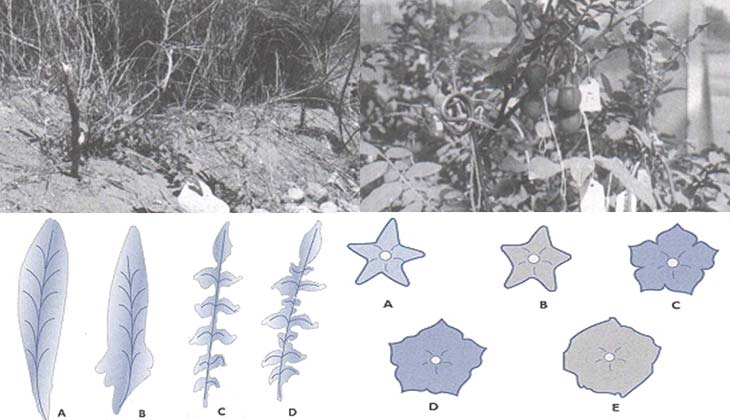
Las papas silvestres crecen en ambientes muy diversos, ubicados entre las cadenas montañosas del sudoeste de los Estados Unidos y el sur de Chile y de la Argentina, pasando por México y América Central. En la Argentina se las encuentra en una vasta región que se extiende desde las provincias del noroeste hasta el sudeste de la de Buenos Aires. Pueden vegetar en las altas montañas (en alturas de hasta 4100m), en valles, quebradas, selvas o costas, y en suelos pedregosos, arenosos, volcánicos o ricos en materia orgánica. Crecen tanto en los ambientes de elevada humedad de las áreas tropicales y subtropicales como en zonas desérticas en las que el agua sólo proviene de nieblas costeras.
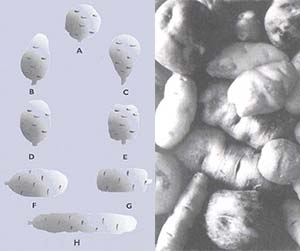
Su exitosa colonización de medios muy distintos se expresa en la gran variedad de formas que pueden adquirir las especies silvestres (Figs. 1 a 6). Las hay con apariencia muy semejante a la cultivada; pueden tener hojas compuestas por varios foliolos u hojas simples; pueden ser arrosetadas (con hojas muy juntas, como los pétalos de una rosa) y con un tallo mínimo, o medir más de un metro de altura. Las flores pueden presentarse en racimos o ser solitarias sus colores variar del blanco, crema o celeste al rojo, púrpura o tonalidades intermedias. La mayoria de las especies produce tubérculos con el interior blanco, crema o amarillo, pero tambien se los encuentra rojos, violetas.
O púrpuras, con la cáscara o piel de color crema, amarillo, naranja o castaño, pigmentada con rojo o púrpura en toda su extensión o en la zona de los nudos donde nacen los brotes (llamados comúnmente ojos). En gran medida, la variabilidad de las distintas especies de papa está controlada genéticamente y, por lo tanto, es heredable. Debido a ello, el estudio de los materiales que contienen los genes (por lo general llamado germoplasma) es de utilidad en investigaciones básicas y aplicadas de áreas que incluyen, entre otras, la genética, la citogenética y el mejoramiento genético, la bioquímica y la patología.
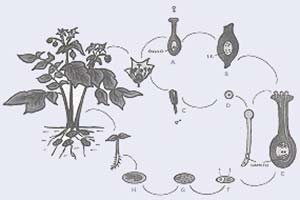
La papa puede reproducirse en forma sexual o asexual. El primer caso (Fig. 7) requiere la participación de los órganos reproductivos femeninos (pistilos) y masculinos (anteras) de las flores, en las que tiene lugar la unión del material genético de los progenitores, que da origen a plantas con nuevas combinaciones de genes. Las generadas por reproducción sexual crecen a partir de semillas que se desarrollan en frutos carnosos (o bayas), semejantes a los del tomate pero de menor tamaño. En la reproducción asexual -llamada también vegetativa- la nueva planta se forma a partir de los tubérculos y, por no haber mezcla de material hereditario, posee la combinación de genes del individuo al que pertenecían esos tubérculos. Esta forma de reproducción da lugar a descendientes genéticamente idénticos a la planta inicial, es decir, a clones de ella.
Durante miles de años, las especies silvestres de papa han estado sometidas a selección natural en condiciones diversas y muchas veces desfavorables. Por eso, muchas han desarrollado características hereditarias valiosas para su supervivencia, que incluyen resistencia a agentes biológicos adversos -insectos, virus, nematodes (gusanos no segmentados de sección circular, algunas de cuyas especies parasitan a vegetales y animales, incluyendo al hombre), bacterias y hongos- y a factores desfavorables no biológicos, como el frío, el calor o la falta de agua. Entre las propiedades hereditarias de las especies silvestres también se encuentran atributos provechosos para su calidad culinaria o industrial, como un alto contenido de materia seca, proteínas y vitaminas. Cuando intervienen en la cruza padres de diferente origen, se produce en la descendencia el fenómeno conocido como vigor híbrido, que se manifiesta como un aumento en el número, tamaño y peso de los tubérculos, comparados con los de plantas cuyos padres sean del mismo origen.
Los cultivos modernos de papa proceden de unos pocos clones llevados a Europa desde el continente americano en el siglo pasado. Por eso, su acervo de genes diferentes -que suele llamarse base genética- es poco abundante. Ello los hace vulnerables a agentes patógenos nuevos o a variantes de los existentes y reduce la diversidad de los ambientes aptos para implantarlos. Un ejemplo dramático de esa vulnerabilidad debida a cultivar grandes extensiones con una única especie -o con especies genéticamente semejantes- fue el ataque de tizón tardío que destruyó los cultivos de papa en Irlanda en los años 1845, 46 y 48. Causado por el hongo Phytophtora infestans, ocasionó una hambruna que resultó en un millón y medio de muertos, determinó la emigración de otro millón de personas y tuvo efectos sociales, políticos y económicos que aún hoy perduran (véase ‘A ciento cincuenta años de la gran hambruna en Irlanda’).
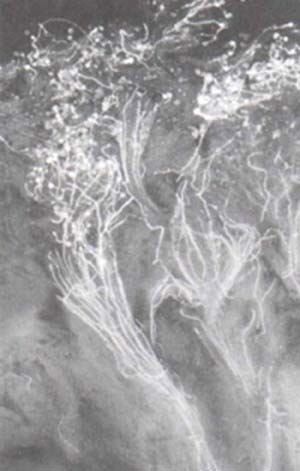
Lo dicho explica por qué los fitomejoradores, cuya tarea consiste en el perfeccionamiento de especies vegetales, han reconocido desde hace años la necesidad de ampliar la base genética de la papa cruzándola con especies silvestres y con cultivos primitivos (los clones plantados por campesinos de los Andes, incluyendo los del noroeste argentino). En muchos casos, es difícil obtener híbridos de papas silvestres y cultivadas, debido a barreras que impiden la unión de las células reproductivas femeninas y masculinas (llamadas gametos) de ambos tipos de especies. Para que esta se produzca, es menester que los granos de polen depositados por insectos o por el viento en la parte superior (o estigma) de los pistilos lleguen a los óvulos, donde ocurre la fecundación, la que sólo sucede cuando hay correspondencia entre la información genética que llevan el polen y el pistilo. De lo contrario, cada etapa de ese avance del polen puede actuar como un obstáculo que impide el cruzamiento (Fig. 8).
Las mencionadas barreras a la hibridación dan lugar a que falle la germinación de los granos de polen, o a que no crezcan los tubos polínicos de una especie en los pistilos de la otra, fenómeno conocido por incompatibilidad cruzada, para diferenciarlo de la incompatibilidad que ocurre entre plantas de la misma especie. Pero aún cuando exista compatibilidad entre el polen y el pistilo y tenga lugar la fecundación, el desarrollo del endosperma o tejido de reserva de la semilla híbrida puede ser anormal e impedir que cumpla con su función nutritiva, lo que causa el aborto del embrión. Tal fenómeno, frecuente en la hibridación de las papas, generalmente se debe a un desequilibrio de genes en el endosperma, que se produce cuando se cruzan plantas con distinto número de cromosomas. Ello afecta al endosperma, porque este se origina igual que el embrión, por la unión de una célula del grano de polen con otra del saco embrionario del óvulo.
Lo mismo que en los animales superiores (véase ‘Cómo se construye un organismo animal’, Ciencia Hoy, 31), la mayoría de las papas silvestres (un 70%) son diploides, esto es, tienen dos juegos de cromosomas (cada uno con doce de estos). La papa cultivada, en cambio, es tetraploide, ya que sus células cuentan con cuatro juegos de ellos (cada uno, también, con doce cromosomas). Dado que, en la mayoría de los cruzamientos entre diploides y tetraploides de papa, el endosperma resulta genéticamente desequilibrado (tiene más genes de un progenitor que del otro), las especies silvestres diploides, cruzadas directamente con papa cultivada son incapaces de producir descendencia.
Para superar estos inconvenientes, en algunos programas de mejoramiento genético, tanto en la Argentina como en el extranjero, se aplican rutinariamente métodos que intentan aprovechar al máximo las posibilidades de la papa cultivada. Tales métodos utilizan la reproducción sexual para obtener nuevas combinaciones genéticas y la asexual para multiplicar las mejores de ellas; también reducen de cuatro a dos el número de juegos de cromosomas de las especies tetraploides, con el fin de obtener descendencia mediante su cruce con diploides e incorporar características agronómicas deseables de estas, pero luego duplican el número de juegos de cromosomas de las plantas hibridas y restauran así la condición tetraploide, óptima para la producción comercial, porque está asociada con los mayores rendimientos de tubérculos (véase ‘¿Cómo se incorpora germoplasma silvestre o de cultivares primitivos en la papa común?’).
Así, manipulaciones realizadas in vivo e in vitro han permitido introducir genes de interés agronómico, provenientes de especies silvestres y de cultivos primitivos, en cultivos comerciales de papa. Entre 1954 y 1986, se crearon en Europa noventa y ocho formas cultivadas (llamadas en jerga cultivares) con genes que confieren resistencia a enfermedades y plagas, y que provienen de diez especies silvestres diferentes. En América del norte se obtuvieron ciento veinticuatro cultivares con germoplasma de once especies silvestres. Por ejemplo, el cultivar canadiense Conestoga tiene genes de dos especies cultivadas y de cinco silvestres, y algunos cultivares europeos poseen genes de hasta seis especies silvestres. El 37% de los cultivares holandeses y el 60% de los alemanes llevan un gen de resistencia a nematodes (denominado Hl), proveniente de una especie oriunda de los Andes, Solanum tuberosum subespecie andigena. La especie Solanum acaule, que crece desde el centro del Perú hasta el noroeste de la Argentina, posee un gen que le confiere resistencia al virus X de la papa (denominado Rxacl); ese gen ha sido introducido en la papa cultivada, mediante procedimientos de mejoramiento genético: por eso está presente en varios cultivares canadienses, americanos y europeos, y en el argentino Serrana INTA. Genes de Solanum tuberosum subespecie andigena, Solanum demissum y Solanum tuberosum subespecie phureja integran la dotación genética de casi todos los cultivares europeos.
Debe señalarse que los métodos que utilizan la reproducción sexual para acceder a parte de la inmensa riqueza genética de las especies silvestres de papa también tienen ciertas desventajas. Junto con las características agronómicas deseables, se transmiten otras que no lo son, como ojos profundos, tubérculos muy pequeños y numerosos, carne pigmentada, etc., que deben ser eliminadas en pasos posteriores del mejoramiento genético, en los que se pierde parcialmente el vigor híbrido logrado en los cruces originales. Por ello, en el futuro los nuevos cultivares posiblemente se basen en construir previamente, de manera racional, un genotipo deseado (se llama genotipo a la constitución genética del individuo). Si, por ejemplo, cultivares de alto rendimiento y bien adaptados a distintas áreas geográficas presentaran ciertas falencias, como resultar susceptibles a determinados agentes patógenos, se les podria transferir genes de especies silvestres y de cultivares primitivos que les confirieran la necesaria resistencia, pero no mediante los procedimientos de cruce que fueron mencionados sino por técnicas de ingeniería genética, las que permitirían mantener el genotipo del cultivar, pero agregarle las nuevas cualidades. La papa se adapta muy bien a este tipo de manipulaciones. De hecho, se han introducido genes de resistencia a virus, heladas y otros factores, en cultivares de los Estados Unidos y de varios países de Europa y de América latina, algunos de los cuales ya están en el comercio. Pero debe tenerse en cuenta que introducir genes por ingeniería genética no conduce a la variabilidad que se logra mediante la reproducción sexual. Esta genera suficientes combinaciones de caracteres heredables como para que comprendan algunos eventualmente útiles ante las situaciones cambiantes del ambiente y de los mercados.




