




En niños de familias carenciadas se observan caracteres sexuales secundarios que se alejan de lo normal. Este artículo explica las causas de tal fenómeno.
La desnutrición, aun cuando actúe por igual sobre ambos sexos, puede alterar la manifestación de algunos caracteres sexuales secundarios.

Las diferencias morfológicas, fisiológicas, bioquímicas y de comportamiento de machos y hembras se denominan, en su conjunto, dimorfismo sexual. Cuando los rasgos físicos diferenciales, o rasgos dimórficos, están directamente vinculados con la reproducción como es el caso de testículos, ovarios, hormonas gonadales, etc. se los denomina caracteres sexuales primarios, mientras que otras notorias diferencias entre los sexos, los caracteres sexuales secundarios que se relacionan con el tamaño corporal, el desarrollo muscular y esquelético, la distribución del tejido adiposo subcutáneo, etc. no están ligados a la función reproductora (véase recuadro “Herencia y caracteres sexuales primarios”)
Aunque la clasificación de los caracteres sexuales en primarios y secundarios es útil y parece inobjetable, existen situaciones en las que no resulta fácil establecer si se trata de unos u otros. Hace ya más de un siglo, Darwin planteó este problema en The Descent of Man and Selectian in Relation to Sex, Londres, 1889, y lo resolvió como era de esperar desde una perspectiva dinámica: mientras los caracteres sexuales primarios serian producto de la selección natural, los secundarios resultarían de la selección sexual (véase recuadro “Selección natural y selección sexual”).
Decir que un carácter sexual es producto de la selección sexual puede parecer redundante, pero ocurre que existen rasgos dimórficos que no están vinculados con la reproducción. En esta categoría se ubican tanto los rasgas neutros como aquellos asociados con alguna actividad ajena a la reproducción. Las diferencias de coloración existentes en ciertos animales que no pueden distinguir los colores, las varias formas relacionadas con el hermafroditismo y las diferencias de comportamiento de algunos invertebrados que no pueden desplazarse constituyen ejemplos de carácter sexual neutro. Sin embargo. este dimorfismo neutro debe integrar el conjunto de caracteres sexuales secundarios, porque todos, de una forma u otra, son seleccionados durante la evolución. La selección natural es también responsable del polimorfismo sexual, o sea, las diferencias de caracteres secundarios en un mismo sexo. En estos casos actúa la llamada selección diversificante, que incrementa la variabilidad de la población y favorece la aparición de grupos de individuos de un mismo sexo con caracteres propios.
La concepción darwiniana del dimorfismo sexual es producto de una paciente y rigurosa observación de la naturaleza; está también sorprendentemente cerca de la interpretación actual de este problema. En la época de Darwin ni siquiera se conocían las leyes básicas de la herencia, pues, a pesar de que contemporáneamente vivía Mendel, los resultados de los experimentos de este sólo fueron ampliamente difundidos a principios del siglo XX, por lo que Darwin no los conoció.
Sin el auxilio de la genética era imposible saber la verdadera causa de la variación aleatoria y, por lo tanto, no se podian considerar los procesos de diferenciación el sexual incluido como producto de una interacción genéticoambiental. El fenotipo de cada individuo el conjunto de sus características observables es producto de su herencia genética y de la acción modificadora del ambiente. Hoy, esta acción ambiental reside, por ejemplo, en el cambio de hábitos alimentarios, la contaminación creciente, malnutriciones de diverso tipo e intensidad y las situaciones de tensión que encontramos en la sociedad moderna, cada vez más competitiva y compleja. Si la medición de un parámetro corporal reflejara directamente la interacción genéticoambiental, se podría evaluar la influencia de cada uno de los factores mencionados. Pero simples comparaciones estadisticas no permiten extraer conclusiones de ningún tipo; es necesario realizar otra clase de estudios. Entre las aproximaciones experimentales accesibles al investigador se cuentan las de genética cuantitativa, que pueden distinguir entre la influencia genética y la del ambiente. También cabe realizar experimentos, de tal forma que se analice el efecto particular de cada factor; las observaciones obtenidas se vuelcan luego a un modelo general. En resumen, podríamos afirmar que la concepción evolutiva moderna permite concebir el dimorfismo sexual como un fenómeno de población, resultante de una interacción genéticoambiental factible de ser estudiada sobre la base de comparar ciertos caracteres, observaciones que luego deben ser corroboradas en forma experimental.
Sabemos que la mayoría de los conceptos evolutivos básicos son aplicables a los seres humanos, con la diferencia de que las poblaciones humanas transmiten también patrones culturales de conducta. Tal transmisión, que se realiza durante muchas generaciones, obedece a mecanismos diferentes de los de la herencia natural, pues los humanos, debido a los factores socioculturales, alteramos nuestro ambiente físico y, en consecuencia, nuestros propios patrones evolutivos. En la adaptación humana no sólo intervienen factores del medio natural; se trata de un complejo proceso que nos lleva al concepto de gran ambiente. Esto implica considerar como factores del medio a un conjunto heterogéneo de fenómenos, desde prácticas consagradas por la costumbre, que se remontan a la noche de los tiempos y que aún perviven hasta lo inverosímil, hasta complejas relaciones de la sociedad moderna que, en forma no menos inverosímil, justifican la existencia, en un mismo tiempo y lugar, tanto de la opulencia de los que pueden comer sin alimentarse piénsese en nuestros mercados atiborrados de menús light y productos diet como de la miseria de los que deben alimentarse deficientemente por falta de recursos económicos. Las condiciones socioeconómicas adversas que soportan los segundos van configurando subpoblaciones sometidas a condiciones adaptativas propias de un ambiente nutricionalmente deficitario.
Estas subpoblaciones permanecen inmersas en la población mayor, pero sus caracteristicas se acentúan mientras perdura la acción nociva del medio y se trasladan por vía no genética a la descendencia. Un niño que nace en un medio económicamente pobre se verá obligado a desarrollar hábitos y comportamientos que le permitan sobrevivir en ese medio adverso, hábitos que atentarán contra una educación eficiente y contra su normal aptitud para competir en la obtención de empleos y otros medios que, llegado a edad adulta, le permitirian mejorar su posición. Por otro lado, una persona que nace en un ambiente de abundancia e hipernutrición parece orientada a cuestiones de otro orden, como adoptar la última moda en materia de regimenes dietéticos tan sólo para acceder o conservar una forma corporal acorde con los criterios estéticos de su tiempo y estrato social. Estas manifestaciones extremas de comportamiento social son producto de una divergencia progresiva de miembros de una misma sociedad, y podrían configurar un caso particular de evolución diversificante. Las curvas de crecimiento de varones y niñas pertenecientes a los llamados bolsones de pobreza no sólo evidencian valores más bajos que los normales sino, también, presentan una nueva relación de equilibrio entre los sexos. Aun en ambientes semejantes, la desnutrición de los varones puede ser mayor que la de las mujeres.
Si se considera que los primeros son, por lo general, de mayor tamaño, puede ocurrir una inhibición o, a veces, hasta una inversión del dimorfismo sexual secundario. Esto provoca que las niñas presenten mayor peso y estatura que los varones durante el desarrollo puberal, y que se observe un retraso en el desarrollo corporal de los varones, lo cual se advierte en las respectivas curvas de crecimiento. No todos los órganos y tejidos responden por igual a la desnutrición crónica; resultan más afectados aquellos de desarrollo lento y constante, mientras que los de crecimiento rápido presentan mayor resistencia al cambio.
El perímetro del brazo, por ejemplo (Fig. 1), está influenciado tanto por la cantidad de tejido óseo como de muscular y adiposo. Si se hacen las correcciones correspondientes, puede calcularse la influencia de cada uno de estos tres componentes en niños normales y en desnutridos. Aunque la comparación directa de los valores de los perímetros arroje cifras similares, puede suceder que exista mayor desarrollo muscular, combinado con menor acumulación adiposa, en niños normales, mientras que los carenciados presenten menor desarrollo muscular y mayor reserva relativa de grasa; se trata, por lo tanto, de un movimiento compensatorio que permanece oculto.
La aseveración de que una deficiencia nutricional puede alterar las diferencias entre los caracteres sexuales secundarios o sea, desencadenar una inhibición dimórfica puede resultar algo difícil de entender, ya que la desnutrición actuaría por igual sobre ambos sexos. Sin embargo, sabemos que, ante un factor adverso, las mujeres exhiben mayor resistencia que los varones a inhibir su crecimiento, y que se mantienen en lo posible dentro de sus respectivos canales de normalidad. Sometidos a las mismas condiciones de carencia, los varones muestran una disminución más importante de peso corporal y cambios en su constitución esquelética, reserva adiposa y/o desarrollo muscular.
Veamos cómo la hipótesis anterior la mayor resistencia femenina a un cambio ambiental adverso pudo ser comprobada en la población carenciada de Villa IAPI (véase CIENCIA HOY, 30:41, 1995). En la figura 2 se muestra un resumen de las observaciones realizadas, las que nos llevan a postular la existencia de dos tipos de patrones dimórficos, el temprano y el tardío.
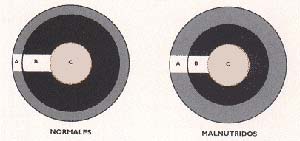
En el patrón dimórfico temprano, la diferencia entre los sexos comienza a manifestarse en etapas prepuberales, como sucede, por ejemplo, con el perímetro craneano y los pliegues subcutáneos tricipital (medido en el músculo tríceps, en la parte posterior del brazo) y subescapular (medido debajo del omóplato). Se observa que, mientras el perímetro craneano es mayor en los varones que en las mujeres, los valores de los pliegues subcutáneos están significativamente disminuidos en los primeros.
El incremento de grasa subcutánea en las mujeres se explica por la necesidad de contar con un reserva energética para futuros embarazos. El patrón dimórfico tardío aparece en la etapa puberal; por ejemplo, a partir de los doce años tanto el peso corporal como la longitud del tronco es menor en los varones que en las mujeres. Otras variables normalmente dimórficas, como la estatura total y el perímetro del brazo, no presentaron dimorfismo sexual. El perímetro craneal fue la única variable que, en Villa API, permaneció normal, es decir, fue mayor en varones que en mujeres, indicio de que la masa encefálica no resultaría afectada por la desnutrición cuando esta se instala después de la lactancia.

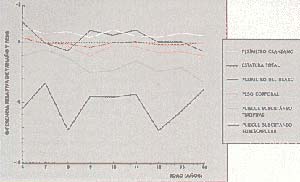
Se observó, además, que la estatura promedio era mayor en las mujeres que en los varones, y que no se producía el cambio hacia el predominio masculino, como debería haber ocurrido antes de los catorce años. Con el propósito de corroborar en forma experimental este tipo de observaciones, realizamos una serie de ensayos con animales. Preparamos dos lotes de ratas albinas de uno y otro sexo: uno para fines de control, que fue alimentado con una dieta normal, y otro que recibió cantidades normales de una dieta pobre en proteínas (lote de ratas desnutridas). Después de cierto tiempo, observamos que el dimorfismo craneano sexual, normal entre animales del grupo control, fue inhibido significativamente en los animales desnutridos, lo cual corroboraba la diferencia entre sexos observada en el ser humano (Fig. 3).
En un trabajo posterior estudiamos el efecto de la castración de los machos sobre el dimorfismo craneano de las ratas, y observamos que este procedimiento producía una inhibición semejante al efecto de la desnutrición (Fig. 4).
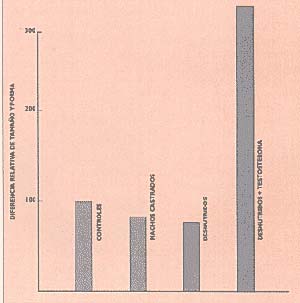
Administramos entonces testosterona la hormona masculina producida por los testículos a ratas macho desnutridas y encontramos que se corregía la inhibición del dimorfismo producida por la malnutrición. Estos resultados experimentales corroboran la hipótesis de que las alteraciones dimórficas observadas en los niños de Villa IAPI sean la resultante de una disfunción de las gónadas masculinas.
La última etapa consistió en estudiar el efecto de otro probable intermediario, la hormona de crecimiento (secretada por la hipófisis, glándula situada debajo del encéfalo) sobre el dimorfismo producido por la malnutrición. Comparamos ratas normales y desnutridas, machos y hembras, con administración de hormona de crecimiento y sin ella, y hallamos que, mientras el dimorfismo en normales no fue influido por la hormona, esta lo restituyó a sus valores normales en los animales desnutridos (Fig. 5).
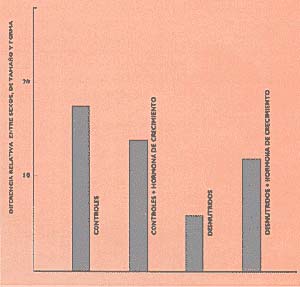
Concluimos que, si bien la disfunción gonadal masculina es responsable directa de la inhibición del dimorfismo craneano producida por la desnutrición, su efecto está relacionado con la actividad de la hipófisis. El tema que hemos abordado no está de ninguna manera agotado. En primer lugar, debe tenerse en cuenta que no sólo las carencias calóricoproteicas afectan el crecimiento; deficiencias en elementos como calcio, zinc y fósforo y aun de algunas vitaminas pueden inhibir el desarrollo psíquico y corporal (véase CIENCIA HOY, 3:14,1991).
En segundo lugar, existen factores socioculturales ligados a un tratamiento discriminatorio de los sexos que, por favorecer primordialmente a los varones en etapas tempranas de su crecimiento, también conducen a una alteración del dimorfismo sexual. De todos modos, la evidencia experimental permite arriesgar una generalización acerca de la dinámica del dimorfismo sexual: si un hecho observado en el ser humano es experimentalmente corroborado en otro mamífero, existe fundamento biológico para inferir la existencia de un modelo general de comportamiento válido para todos los seres de, por lo menos, el mismo nivel taxonómico.




