




Estamos rodeados de microorganismos patógenos, como virus, bacterias, hongos y parásitos. A pesar de tal exposición continua a agentes agresores, rara vez padecemos de infecciones graves. ¿Cómo hace el organismo para defenderse?
Este artículo está principalmente orientado a actualizar los conocimientos de quienes han tenido contacto con el tema pero desconocen sus avances más recientes. Sugerimos al lector poco familiarizado con la biología actual que, después de leer el primer párrafo, vaya al recuadro ‘Un poco de historia’ y luego consulte puntualmente los sucesivos recuadros a medida que el texto remita a ellos.
El término inmunidad deriva del latín immunitas, el privilegio en virtud del cual los senadores romanos estaban exentos de ciertos deberes cívicos. En las ciencias biológicas, inmunidad significa protección contra enfermedades, en especial, las infecciosas.
Los vertebrados –entre ellos el hombre– han desarrollado mecanismos para protegerse de las agresiones que les vienen del ambiente, causadas por bacterias, virus, parásitos y toxinas, entre otros agentes (los que reciben el nombre de patógenos, o causantes de enfermedad.
Algunos de esos mecanismos son de tipo general y constituyen una primera línea de defensa del organismo. Hablamos, así, de barreras naturales –la piel, las mucosas– y de inmunidad innata o inespecífica; de esta clase, por ejemplo, es la fagocitosis (de jagein –phageîn–, comer, y kutoV –kytos, célula), proceso por el cual ciertas células denominadas macrófagos, que se encuentran en la mayoría de los tejidos, capturan y digieren microorganismos. Decimos que esta defensa es innata porque no depende de la exposición previa al patógeno. Pero en todos los vertebrados puede activarse también una segunda línea de defensa, un sistema de reconocimiento específico del agente agresor. Tal mecanismo se denomina sistema inmune y está constituido por dos tipos de células:
(a) las células accesorias, como son los macrófagos y las células dendríticas (de dendron –déndron–, árbol, así llamadas porque poseen ramificaciones), que participan en la captura y degradación (también denominada procesado) del antígeno, y
(b) las células inmunocompetentes o linfocitos, que pueden reconocer específicamente tanto al antígeno nativo como al procesado por las células accesorias.
Las células del sistema inmune se originan durante la hematopoyesis, el proceso de formación de células sanguíneas, cuyas características les permiten cumplir funciones vitales específicas. Así, los eritrocitos o glóbulos rojos transportan oxígeno, las plaquetas promueven la coagulación de la sangre para evitar hemorragias y las células blancas o leucocitos (que incluyen a los linfocitos, los monocitos y los neutrófilos), junto con los macrófagos, forman el ejército celular del sistema inmunológico. Todas derivan de las células hematopoyéticas troncales, también denominadas células indiferenciadas multipotentes porque pueden replicarse indefinidamente y diferenciarse en varias clases de células secundarias. Estas, las precursoras, a su vez, originan líneas celulares diferenciadas (por ejemplo, los eritrocitos). Las células hematopoyéticas troncales aparecen primero en el saco amarillo embrionario, pero, a medida que el feto se desarrolla, migran al hígado. Inmediatamente después del nacimiento, la hematopoyesis pasa a ser función de la médula ósea.
Las distintas estirpes de linfocitos constituyen entre el 1% y el 10% de las células que produce diariamente el cuerpo; se encuentran principalmente en la superficie de las mucosas y en los órganos linfoides, nombre que se aplica a la médula ósea, al timo (localizado en la parte superior del pecho, detrás del esternón) y a otros órganos llamados periféricos (los ganglios o nódulos linfáticos, el bazo y ciertos tejidos asociados con el intestino, como las placas de Peyer). Existen poblaciones de linfocitos que cumplen diferentes funciones, relacionadas con el órgano linfoide en el que maduraron (o adquirieron competencia inmunológica). Los linfocitos B, productores de anticuerpos, maduran en la médula ósea (o en un órgano equivalente de las aves, denominado bursa de Fabricius, de donde proviene la denominación de linfocito B). Los linfocitos T lo hacen en el timo, donde se diferencian en linfocitos T citotóxicos (Tc), que destruyen las células infectadas, y linfocitos T colaboradores (Th, por helper), que ayudan a los B y a los macrófagos a cumplir su función.
Durante el proceso de maduración, el contacto de los linfocitos B y T con células epiteliales de los órganos linfoides respectivos les confiere competencia inmunológica; es decir, hace que adquieran una función inmunológica específica. Sin embargo, aunque cumplan diferentes funciones, las poblaciones linfocitarias son indiferenciables desde el punto de vista morfológico.
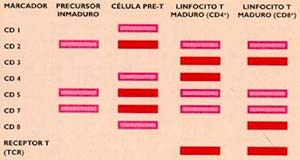
El linfocito B pasa por sucesivos estadios de diferenciación, conocidos como célula Pro-B o precursor inmaduro, célula Pre-B,linfocito B inmaduro y linfocito B maduro; este, a su vez, se diferencia en célula plasmática o plasmocito, la célula que secreta anticuerpos. Durante esos estadios aparecen diferentes moléculas en la membrana celular que constituyen indicadores (o marcadores) específicos y permiten establecer el grado de madurez de las células (Fig.1). Algunas de esas moléculas –que han podido ser identificadas mediante el uso de anticuerpos monoclonales y forman parte del grupo denominado cluster o conjuntos de diferenciación (CD) perduran durante la mayor parte de la diferenciación del linfocito, en tanto que otras son propias de ciertos estadios (véase el recuadro ‘Identificación de antígenos celulares’).

El marcador más significativo del linfocito B es la presencia en la membrana de una molécula de inmunoglobulina de la clas e M, o IgM (véase el recuadro ‘Los anticuerpos’), que reconoce e antígeno; por ello, esa IgM del linfocito B es denominada, también, receptor antigénico y tiene la misma especificidad –es decir, reconoce el mismo sitio antigénico– que el anticuerpo que será secretado luego de la estimulación de la célula por el impacto del antígeno (véanse los recuadros ‘Los receptores de los linfocitos’ y ‘La transducción de señales’).
Si células indiferenciadas, al migrar por el cuerpo, encuentran el microambiente del timo, se diferencian en linfocitos T. Los primeros cambios que sufren como parte de ese proceso de maduración ocurren en la corteza de dicho órgano, donde los linfocitos desarrollan en sus membranas (se dice también que las células expresan una molécula, cuando esta se detecta en la membrana celular) moléculas pertenecientes a los conjunt os de diferenciación (CD2, CD5 y CD7 en la figura 2). La etapa siguiente tiene lugar en la médula del timo, donde se incorporan nuevas moléculas a la superficie celular, entre ellas, las correspondientes a los CD1, CD4 y C D8. Después, el linfocito T pierde algunas de esas moléculas de superficie e incorpora otras, como las del CD3. Para entonces, las células se han diferenciado en dos grupos: el que expresa CD4 y el que expresa CD8, respectiva mente marcadores de los linfocitos Th y Tc, como lo muestra la misma figura 2. En estas etapas los linfocitos T reordenan los genes que permiten sintetizar su receptor antigénico, un heterodímero formado por dos cadenas peptídicas, a/b o g/d (vuélvase al recuadro ‘Los receptores de los linfocitos’). Cuando las células completan su maduración se vuelcan al torrente sanguíneo como linfocitos T maduros, en dos grupos definidos, los CD4+/CD8- y los CD8+/CD4-
Los Anticuerpos
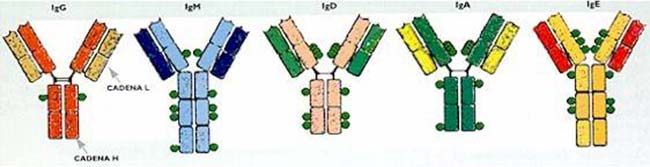
Con el nombre anticuerpos se designa a unas proteínas que se encuentran en el plasma –la parte fluida de la sangre– y en los fluidos extracelulares. Como los líquidos del organismo alguna vez fueron llamados humores, se dice que los anticuerpos son el instrumento de la inmunidad humoral. Forman parte de la familia de las inmunoglobulinas (Ig), que son moléculas con forma de Y, en cada uno de cuyos brazos superiores se encuentra un sitio que permite la unión con un antígeno (véase ‘La estructura básica de un anticuerpo’, Ciencia Hoy, 11:38-39, 1991).
La estructura química de los sitios del anticuerpo que pueden adherirse al antígeno son diferentes para cada molécula de anticuerpo, lo que confiere enorme diversidad a la respuesta inmune, pues siempre habrá un anticuerpo que pueda unirse a uno de los tantos antígenos que nos rodean (véase ‘Cómo se origina la diversidad de los anticuerpos’, Ciencia Hoy, 11:41, 1991). Tales sitios no reconocen bacterias enteras, sino zonas relativamente pequeñas de estas –llamadas epitopes odeterminantes antigénicos–. Cada sito de unión del anticuerpo puede adherirse a un único epitope. La unión entre el anticuerpo y su ntígeno se produce porque las conformaciones moleculares de ambos son complementarias y se atraen. Varias fuerzas estabilizan esa unión, entre ellas la atracción que existe entre cargas eléctricas negativas y positivas y la capacidad de ciertos grupos químicos de interactuar eliminando el agua de la interfase (uniones hidrofóbicas). La estructura química del pie de la Y de las moléculas de inmunoglobulina, contrariamente a sus brazos, no varía para cada una de aquellas, por lo que se denomina la región constante del anticuerpo. Adopta por lo general una de cinco formas más importantes, o isotipos, que determinan la clase de inmunoglobulina y su funcionalidad (Figura).
Los anticuerpos se unen a los agentes patógenos o antígenos en el espacio extracelular, y aseguran la protección del organismo mediante tres mecanismos:
(a) pueden neutralizar al patógeno o a sus productos tóxicos adhiriéndose a ellos e impidiendo la infección o la toxicidad;
(b) pueden facilitar la captura de los patógenos por las células fagocíticas mediante un proceso denominado opsonización, que altera la membrana del cuerpo extraño, y
(c) pueden activar el sistema de complemento, constituido por una serie de proteínas plasmáticas que ayudan a los fagocitos a ingerir y destruir las bacterias. Todos los patógenos y partículas extrañas unidas a anticuerpos finalizarán en poder de los fagocitos, que los destruirán. El sistema de complemento y los fagocitos no reconocen al antígeno: son los anticuerpos los que les señalan su existencia.
Los linfocitos CD4+ o T colaboradores (Th) se diferencian, a su vez, en dos poblaciones funcionalmente distintas: Th1 y Th2. La primera sintetiza y libera las citoquinas denominadas interleuquina 2 e interferón g (véase el recuadro ‘Las citoquinas’) y estimula a los macrófagos a degradar bacterias y parásitos que se replican en las vesículas intracelulares (véase el recuadro ‘Los compartimentos celulares’). Los Th2, que producen las interleuquinas 4, 5 y 10, cumplen funciones regulatorias, como es la cooperación con los linfocitos B.
Los linfocitos CD8+ son los citotóxicos (Tc), que destruyen las células infectadas con, por ejemplo, virus; además de secretar interleuquina 10 e interferón g, liberan enzimas proteolíticas (así llamadas porque de gradan las proteínas) y contienen gránulos secretorios de perforina, una proteína que forma canales (por donde se escapa el material celular ) en las membranas de las células atacadas,con la consiguiente destrucción de estas.
Los linfocitos B y T que adquieren competencia inmunológica van a poblar los órganos linfoides secundarios, aunque también circulan por la sangre. Los aproximadamente mil millones de linfocitos que tiene un adulto normal, cada uno con un receptor antigénico diferente en su membrana, resultan suficientes para proteger a una persona de los agresores externos, pues se calcula que el número de antígenos existentes en la naturaleza oscila entre diez y cien millones.
Compartimientos Celulares
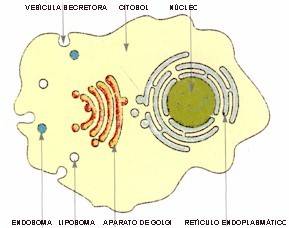
En una célula hay dos compartimentos principales, separados por membranas. Uno es el citosol, que se continúa en el núcleo de la célula por los poros de la membrana nuclear; el otro es el sistema vesicular, que comprende el retículo endoplásmico, el aparato de Golgi, los endosomas, los lisosomas y otras vesículas intracelulares (Fig). El sistema vesicular está en estrecho contacto con el espacio extracelular. Así, las vesículas secretorias que se forman en el retículo endoplásmico (donde se acumulan las proteínas sintetizadas por la célula) son transportadas por el Golgi y descargan su contenido fuera de la célula o en su membrana, mientras los endosomas introducen el material extracelular en el interior del sistema vesicular.
El linfocito T reconoce Pequeños péptidos de siete a quince aminoácidos (provenientes del procesado del antígeno por las células presentadoras) asociados con las moléculas HLA presentes en las membranas (véase el recuadro Los antígenos de histocompatibilidad). Cuando los antígenos son sintetizados dentro de la célula, como es el caso de las proteínas virales, los péptidos se unen con las moléculas HLA de clase I, en tanto que los antígenos de síntesis exógena (bacterias, parásitos, proteínas extrañas), previa endocitosis y procesado, son presentados al linfocito T por los HLA de clase II. La capacidad de los infocitos T maduros para reconocer el elemento restrictivo. las moléculas HLA-I o HLA-II propias no se hereda: se adquiere durante la maduración linfocitaria, por la cual los linfocitos T, por un mecanismo de doble selección positiva y negativa, aprenden a reconocer los complejos formado s por las moléculas HLA y los péptidos procedentes de la degradación de antígenos extraños, así como los complejos formados con péptidos provenientes de tejidos propios. Sólo el 5% de las células que se diferencian en el timo sobreviven y pasan a la circulación como linfocitos maduros.
Las que mueren durante el proceso de selección lo hacen por apoptosis, o muerte celular programada, inducida por señales celulares específicas (véase ‘El suicidio celular’, Ciencia Hoy, 11:43, 1991), proceso que evita que proliferen los linfocitos T que reconocen moléculas endógenas. Si el mecanismo fallara, se originaría una respuesta inmune dirigida contra los tejidos y células propios y se produciría una enfermedad autoinmune o de autoagresión.
Los Receptores de los linfocitos
El linfocito B reconoce al antígeno por medio de su receptor antigénico, una inmunoglobulina M de membrana, así denominada por ser un anticuerpo que, en lugar de circular por la sangre, está anclado en la membrana celular (Figura). Existen otras dos proteínas asociadas con el receptor antigénico, la Iga y la Igb, que se encuentran unidas entre ellas en forma covalente. En 1976, estudiando los cromosomas de linfocitos inmaduros, Susumo Tonegawa encontró que los genes que llevan la información mediante la cual se sintetizan los anticuerpos están muy alejados unos de otros, mientras en las células plasmáticas, productoras de anticuerpos, se hallan estrechamente asociados.En otras palabras, se produce un reordenamiento de genes, que iene lugar al azar, durante el proceso de diferenciación de los linfocitos B, por el cual cada uno de estos lleva en su membrana una IgM diferente. Lo anterior da origen a un repertorio de linfocitos del orden de los miles de millones, que se encuentran en espera, por así decirlo, del a llegada de un patógeno. Cuando un antígeno con una estructura complementaria a la de la IgM de la superficie celular impacta al receptor del linfocito B, este se activa y prolifera. Aquí volvemos a encontrar la base molecular de la selección clonal a la que se hizo referencia.
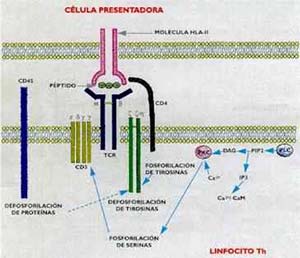
El receptor del linfocito T (TCR, por T cell receptor) está compuesto por dos proteínas variables (cadenas a y b en la figura II), llamadas polimórficas porque la secuencia de sus aminoácidos difiere de un linfocito T a otro; estas moléculas se originan a partir de un reordenamiento de genes semejante al descripto para el receptor del linfocito B. El TCR está asociado al complejo CD3, formado por las cadenas polipeptídicas g, d y e, y a otras dos proteínas, unidas entre sí, que pueden ser iguales en el 90% de las células (cadena z de la figura II) o diferentes en el 10% restante (cadenas z y h de la misma figura). Estas molécula s tienen una estructura similar en todas las células T y participan en los mecanismos de activación celular. Los antígenos celulares CD4 y CD8, que identifican, respectivamente, a poblaciones de células T colaboradoras y citotóxicas, se denominan correceptores. Existen otras proteínas de membrana, denominadas moléculas de adhesión, que se encuentran tanto en las células presentadoras de antígenos como en los linfocitos; su interacción aumenta la afinidad de la unión entre células y da lugar a coestimulaciones fundamentales para la activación de los linfocitos.
La Trasducción de Señales
Denominamos transducción de señales al proceso que ocurre dentro de una célula luego de que esta recibe una señal o indicación externa. Tal señal, en la mayoría de los casos, se produce cuando una molécula (o ligando) se une específicamente con un receptor presente en la membrana celular. El efecto del ligando es desencadenar una cascada de sucesos –apertura de canales iónicos, activación de enzimas y de genes, etc.– que permiten que la célula se active, prolifere, comience a secretar algún compuesto, se diferencie en otro tipo celular, etc.
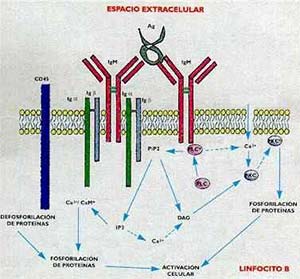
La activación del linfocito B se produce cuando el antígeno se une con el receptor, es decir, con la inmunoglobulina M de membrana.
El antígeno, que hace de puente entre dos IgM, induce la aparición de cierto número de proteínas fosforiladas (es decir, unidas a uno o varios grupos fosfato). Esta fosforilación se debe a la activación de diferentes tirosín quinasas, las enzimas que catalizan la unión de un fosfato con ciertos aminoácidos de las proteínas, entre ellos tirosinas y serinas (véase ‘La estructura de las proteínas’, Ciencia Hoy, 5:32-33, 1995). Como entre esas proteínas fosforiladas del linfocito B se encuentran las denominadas Iga e Igb, se supone que estas cadenas, que están asociadas al receptor antigénico (Fig. I), deben tener un cometido importante en el mecanismo de activación de la célula.
Se ha encontrado que diferentes familias de tirosín quinasas participan en el proceso de fosforilación. Los componentes de los miembros de una de estas familias tienen dos regiones, o dominios, semejantes, denominados SH2 y SH3. Los SH2 son responsables de la interacción entre la enzima y la proteína a fosforilar, mientras que los SH3 pueden unirse con otras proteínas capaces, a su vez, de transducir señales. La actividad de las tirosín quinasas reside en la región C-terminal de estas enzimas. En ese dominio hay dos tirosinas, ubicadas en las posiciones 394 y 505; la fosforilación de la tirosina 394 por otra quinasa produce la activación de la enzima, mientras que la fosforilación de la 505 la inactiva. Se supone que el último es el estado de la tirosín quinasa en el linfocito B en reposo, y que la agregación del receptor B por el puente formado por el antígeno originaría la remoción del fosfato correspondiente al sitio inhibitorio; el CD45, una proteína de membrana con actividad de fosfatasa (es decir, que remueve los grupos fosfato), puede participar en la regulación de este proceso.
Las quinasas activadas fosforilan también a la fosfolipasa C-g (PLC), la que hidroliza un fosfolípido de la membrana, el fosfatidilinositol difosfato (PIP2), en inositol trifosfato (IP3) y diacilglicerol (DAG), según se aprecia en la figura I. El IP3 estimula la salida de Ca2+ de los reservorios celulares y promueve también su entrada desde el exterior, activando a su vez una quinasa dependiente de Ca2+ y calmodulina (Ca2+/CaM). El Ca2+ y el DAG provocan la translocación de otra quinasa (PKC) a la membrana citoplasmática, donde la enzima altera el transporte de varios iones (Fig. I). Todos estos hechos hacen que el linfocito B progrese hacia una fase del ciclo celular caracterizada por la síntesis de ADN. La célula continúa diferenciándose hasta llegar al estado de célula plasmática, que secreta un anticuerpo con un sitio de unión con el antígeno idéntico al de la IgM que formaba el receptor del linfocito B que se activó.
Durante la activación del linfocito T, cuando el complejo TCR/CD3 interactúa con el correspondiente HLA/péptido, ocurre un mecanismo de fosforilación similar al expuesto para el linfocito B (Fig. II). En una o varias de las subunidades del receptor se advierten cambios conformacionales, con la consecuente activación de las quinasas asociadas, que fosforilan diferentes componentes intracelulares. La cadena z es la única subunidad del TCR/CD3 en la que se han detectado tirosinas fosforiladas; a su vez, la porción citoplasmática de esta cadena puede transducir señales durante el proceso de activación del linfocito.
En forma semejante a lo que ocurre con los linfocitos B, la PLC-g de los linfocitos T activados experimenta una rápida y transitoria fosforilación de sus tirosinas; la enzima activada hidroliza PIP2, y origina IP3 y DAG, que estimulan a la PKC. Esta enzima fosforila una variedad de moléculas, siguiendo la cascada ya indicada en la activación del linfocito B (Fig. II).
Las células accesorias (macrófagos y células dendríticas, principalmente) tienen por función procesar los antígenos y presentar los péptidos resultantes asociados a las moléculas HLA de clase I o II, para su reconocimiento por los linfocitos Tc y Th, respectivamente.
Dichas células, especialmente los macrófagos, sintetizan y secretan diversos factores, como la interleuquina 1 y el factor de necrosis tumoral, los que actúan sobre las células inmunocompetentes. Cuando un patógeno ingresa en el organismo se activa un mecanismo de defensa que se denomina sistema de complemento. No entraremos en el detalle de este complejo sistema, pero diremos que, por ese mecanismo, las bacterias son recubiertas por ciertas proteínas que, a su vez, tienen receptores en las células fagocíticas.
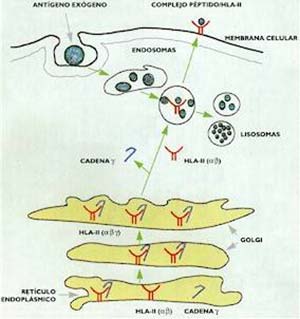
Gracias a ese ‘puente proteico’ el patógeno es incorporado (endocitado) por la célula presentadora y agregado a vesículas los endosomas cuyas enzimas inician su degradación (Fig. 3). El proceso concluye en los lisosomas, también ricos en enzimas proteolíticas, como lo menciona el recuadro ‘Los compartimentos celulares’.
Mientras tanto, en el retículo endoplásmico se realiza el ensamblado de las cadenas a y b de las moléculas HLA-II para formar el dímero a/b, al que se asocia en forma transitoria una tercera cadena peptídica (invariable o g), la que impide su unión con péptidos de origen endógeno. Cuando las moléculas HLA-II llegan al aparato de Golgi, y media una señal no identificada hasta el presente, el complejo a/b/g se dirige a los endosomas que contienen el antígeno procesado; en el camino, la cadena g se degrada y el dímero a/b puede unirse entonces con los péptidos provenientes del procesamiento del antígeno exógeno, como se muestra en la citada figura 3. Los virus se replican dentro de las células y el linfocito T debe reconocer y eliminar las células infectadas para impedir el crecimiento desproporcionado del agente infeccioso.
Los Antígenos de Histocompatibilidad
Cuando se comenzó a injertar órganos en animales, se advirtió la existencia de moléculas en los tejidos que determinaban si el injerto sería rechazado por el hospedante (o huésped); tales moléculas fueron denominadas antígenos de histocompatibilidad (de istoV –histós–, tejido). Ahora sabemos que tales antígenos son moléculas ancladas en las membranas celulares, y que su síntesis está dirigida por un gran número de genes, que se denominan el complejo mayor de histocompatibilidad, o MHC (por Major Histocompatibility Complex). En los humanos existe un equivalente del MHC, que se denomina HLA, por Human Leucocyte Antigens, ya que originalmente fue detectado en las células blancas de la sangre.

La función de las moléculas HLA consiste en unir pequeños péptidos resultantes de la degradación intracelular de los patógenos y llevarlos hacia la membrana celular, donde el complejo péptido/HLA es reconocido por los linfocitos T. Puesto que tal mecanismo es responsable de la destrucción de los patógenos, estos tienden a cambiar su estructura, por mutación de sus genes, para pasar inadvertidos y escapar de ser puestos en presencia del linfocito T. Para que lo último no suceda, el complejo HLA está dotado de dos características importantes. La primera es que existen varios genes que contienen la información (dicho más técnicamente, que codifican ) para que tenga lugar la producción (o síntesis) de las proteínas HLA, las cuales, además, al combinarse, originan diferentes moléculas, por lo que se dice que el complejo es poligénico. La segunda es que también es polimórfico, debido a que en una misma especie hay variaciones de cada gen y, por lo tanto, de su producto. En las especies diploides (que tienen dos juegos de cromosomas), cada gen o sus variantes están presentes dos veces, pues hay una versión en cada cromosoma homólogo; cada miembro de esas duplas de genes se denomina un alelo. Los alelos pueden ser dominantes (cuando la presencia de un solo ejemplar de una variante basta para que se produzca o sintetice la proteína que codifica) o recesivos (cuando dicha proteína no se sintetiza o expresa a menos que estén presentes dos ejemplares de la misma variante). Para el complejo HLA, existen decenas de alelos diferentes que se heredan de a pares (uno del padre y otro de la madre) y se manifiestan simultáneamente (en otras palabras, que son codominantes). El resultado de este arreglo genético es que se sintetizan moléculas polimorfas de HLA, diferentes de individuo en individuo. Co ntrariamente, denominamos monomorfas a las proteínas del complejo HLA que tienen la misma estructura en todos los individuos de una misma especie.
Las moléculas HLA de clase I se encuentran en la membrana de todas las células humanas, excepto los eritrocitos y los espermatozoides, mientras que los HLA de clase II solo se hallan en las células presentadoras de antígenos y en los linfocitos B. El complejo HLA-I está constituido por una cadena polipeptídica a, polimórfica, inserta en la membrana celular, asociada no covalentemente con otra proteína, la b2, una microglobulina monomorfa (Fig. IA). L os HLA-II están formados por dos cadenas peptídicas polimórficas, a y b, insertadas en la membrana celular y asociadas entre ellas en forma no covalente (Fig. IB). En el citoplasma celular y antes de pasar a la membrana, los HLA-II se hallan unidos a una tercera cadena peptídica denominada invariante o cadena g, que no presenta isoformas y desempeña un papel importante en el transporte del péptido antigénico (véase la figura 3 del texto principal).
Tanto en los complejos HLA-I como HLA-II, el plegamiento de las proteínas origina una hendidura donde se ubica el péptido proveniente del patógeno, el que puede estar formado por ocho a diez aminoácidos cuando se une con las moléculas HLA-I, o por más de trece de ellos si lo hace con las HLA-II.
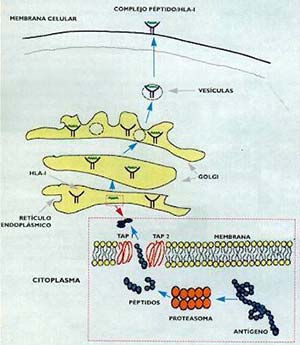
En dichas células se sintetizan las proteínas virales, extrañas y, por lo tanto, antigénicas, que son degradadas en el citosol por los proteasomas, una maquinaria enzimática formada por veintiocho subunidades (Fig. 4). Los péptidos son luego transferidos a las unidades transportadoras (o TAP, por Transporters Associated with Antigen Processing) que se encuentran en la membrana del retículo endoplásmico, las que los conducen hacia el interior de este. Una vez en el Golgi, los péptidos se unen con las moléculas HLA-I y, después, son transportados a la membrana celular para su reconocimiento por el receptor del linfocito T (véase la figura 4).
La unión del receptor del linfocito T con el complejo péptido/HLA no es suficiente para desencadenar una respuesta inmune.
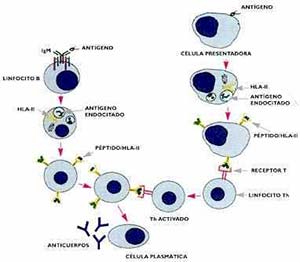
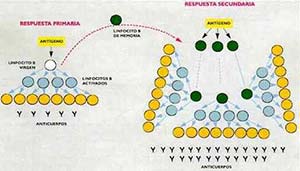
Para que ocurra se necesita que la célula presentadora de antígeno y el linfocito T se adhieran firmemente, lo que ocurre por medio de interacciones entre otras moléculas presentes en las membranas de ambas células, las denominadas moléculas de adhesión. A diferencia de los linfocitos T, los B reconocen antígenos nativos, en solución, y sin necesidad de célula presentadora o restricción HLA (consúltese nuevamente el recuadro ‘Los receptores de los linfocitos’). Sin embargo, para lograr una buena respuesta inmune humoral (o sea, para que se formen anticuerpos) se necesita la cooperación de un linfocito Th. ¿Cómo se explica lo anterior, si el linfocito T reconoce un péptido producto de la degradación antigénica y, en cambio, el receptor del linfocito B reconoce un epitope presente en el antígeno intacto? Se ha podido demostrar que el linfocito B, cuando se ha unido con el antígeno por su receptor, la IgM de membrana, lo endocita y procesa de igual manera que las células presentadoras de antígeno. En otras palabras, el linfocito B también expone en su membrana, asociados a las moléculas HLA- II, pequeños péptidos que son reconocidos por aquellos linfocitos Th que hayan aprendido a hacerlo en la membrana del macrófago (Fig. 5). Como consecuencia de la interacción de las dos células, se incrementa en el linfocito B la cantidad de receptores para interleuquina 2, la cual, sintetizada por el linfocito Th, se une a su receptor y activa la célula B, la que finalmente se diferencia en célula plasmática, productora de anticuerpos. Los hechos que acabamos de describir se ponen en marcha cuando un antígeno se hace presente por primera vez en un vertebrado inmunológicamente maduro. Es la denominada respuesta primaria. El proceso se inicia con unos pocos linfocitos que poseen receptores, los cuales, gracias a su enorme diversidad, pueden reconocer cualquier material extraño: es la selección clonal que menciona el recuadro ‘Un poco de historia’. Pero los linfocitos T y B que no se transforman en células efectoras perduran en el huésped por períodos prolongados como células con memoria inmunológica. Con un nuevo estímulo antigénico tiene lugar la denominada respuesta secundaria, por la que esas células con memoria inmunológica, que, por tener una especificidad definida (ya reconocieron una vez al antígeno) y por integrar una población más numerosa que la primera, forman anticuerpos en mayor cantidad y más rápidamente que en la respuesta primaria (Fig. 6). Este es, en esencia, el mecanismo molecular que explica el éxito de las vacunas: el individuo recibe el patógeno muerto o atenuado y establece una respuesta primaria. Cuando sobreviene la infección, las células con memoria reaccionan rápidamente y eliminan al agente agresor.
Recapitulando, el reconocimiento de un antígeno por las células inmunocompetentes pone en marcha la respuesta inmune. Cuando los linfocitos B y T interaccionan con sus ligandos específicos ocurre una serie de hechos, algunos inmediatos y otros más tardíos, que llevan a la activación de genes y a la síntesis de proteínas reguladoras (vuélvase al recuadro ‘La transducción de señales’). En los linfocitos B hay activación de los genes que permiten la síntesis de los receptores de interleuquina 2 y 4, aparición de moléculas de adhesión, fundamentales para algunas funciones, y aumento de la cantidad de moléculas HLA-II en la membrana celular, lo que facilita la presentación al linfocito T de los péptidos provenientes de la degradación del antígeno. En los linfocitos T se activan los genes de varias interleuquinas y de sus receptores. La aparición del receptor para interleuquina 2 es importante, puesto que esta citoquina, sintetizada por el mismo linfocito T, puede interactuar con su receptor y funcionar como factor de crecimiento sobre la misma célula. Lo descripto, hasta donde sabemos, ocurre durante la activación de los linfocitos B y T y permite entender cómo se pone en marcha el mecanismo, complejo pero eficiente, de la respuesta inmune




