




Una especie está formada por el conjunto de los individuos que -en condiciones naturales- pueden aparearse entre sí; los de especies diferentes no se aparean. Los ejemplares de una misma especie se reconocen mediante señales químicas, acústicas, visuales, táctiles, etc., propias de esa especie.
Los individuos de una misma especie pueden ser distintos entre ellos, pero hay animales de aspecto similar que pertenecen a especies diferentes. ¿Cómo se identifican estas especies gemelas?
Por lo general es sencillo distinguir entre dos animales de especies diferentes: uno puede ser grande y llamativo y el otro pequeño y modesto; aquel amarillo y este negro; el primero tener alas y el segundo no. Tales diferencias groseras son las características que, frecuentemente, utiliza el naturalista para identificar las especies. A veces, sin embargo, pueden resultar engañosas. Por ejemplo, los machos de león o de pavo real son muy distintos de las respectivas hembras, y las hormigas obreras, estériles y ápteras (o sin alas), difieren de los machos y hembras fértiles, generalmente alados.
El taxónomo que se ocupa de clasificar los seres vivos, puede proceder sobre la base de sus observaciones de los animales, las que podrían incluir, entre otras cosas, la constatación al tacto de rugosidades o espinas en la superficie de su cuerpo; si están vivos podría, también, olerlos u oírlos cantar, así como observar su comportamiento, etc. Es usual, sin embargo, que el especialista sólo se enfrente con cráneos, pieles, animales sumergidos en formol o alcohol o especimenes elegantemente ensartados en alfileres. Sea como fuere, en la medida en que el investigador se base en los caracteres morfológicos de los animales para ordenarlos en categorías o taxones, puede no reconocer la existencia de especies gemelas, es decir, especies diferentes formadas por animales de aspecto semejante. Por suerte, los animales parecen saber muy bien cuál individuo es de su misma especie y cuál no (distinguen un coespecífico de un heteroespecífico); en especial, se reconocen en el momento de la reproducción. El grupo de trabajo del autor aprovecha esta circunstancia para identificarlos.
Según el concepto clásico, derivado de la teoría de la evolución, una especie biológica está formada por el conjunto de individuos que en condiciones naturales pueden aparearse entre sí. Esta definición es aplicable, principalmente, a las especies en las que se distinguen machos y hembras, y se basa, sobre todo, en el reconocimiento sexual coespecífico, con el consiguiente rechazo de la copulación heteroespecífica, concepto que denominamos aislamiento reproductor.
Existe en los animales un conjunto de barreras biológicas que evitan el cruzamiento o la hibridación de individuos de distinta especie; son propias del animal y no dependen del medio. Un río, una cordillera o un desierto pueden separar poblaciones, pero, en si mismos, no son mecanismos de aislamiento, como, en cambio, lo son entre otros la preferencia por determinado hábitat (un factor ecológico) o la actividad a distintas horas del día o en diferentes épocas del año (factores vinculados con el comportamiento, o etológicos). En estos casos, las especies quedan efectivamente aisladas unas de otras, pues no coinciden en el espacio o en el tiempo.
¿Cómo se identifican entre ellos los individuos de una misma especie? Desempeñan un papel fundamental ciertas señales químicas, acústicas, visuales, táctiles, etc., características de cada especie. Si no son efectivas, lo que raramente sucede en condiciones naturales, la transferencia espermática puede encontrar dificultades mecánicas, por no ensamblar adecuadamente las estructuras genitales del macho y la hembra.
Sí los mecanismos de aislamiento ecológicos, etológicos y mecánicos son superados, actúan otros postcopulatorios: el esperma o los huevos no sobre viven, o bien resultan engendrados ejemplares híbridos, de baja o nula viabilidad o fecundidad. En los animales actúan, por lo general, varios mecanismos que aseguran el cruzamiento coespecifico; predominan los precopulatorios, los que, por intervenir en forma más precoz, ahorran considerable tiempo y energía. En especial, las hembras tienen mucho que perder por una equivocación y son particularmente selectivas, pues arriesgan una importante cantidad de gametas; podrían malograr una carnada, un período reproductor o toda su descendencia.
Según los conceptos anteriores, la semejanza física entre dos individuos es irrelevante para establecer sí son coespecíficos o heteroespecíficos. En cambio, son importantes características como ser nocturno, reproducirse en otoño, vivir exclusivamente en el bosque serrano, actuar de cierta manera en el cortejo, etc. Es decir, se reconoce la existencia de una especie sí se puede comprobar que para sus integrantes funcionan mecanismos de aislamiento. Efectuado ese reconocimiento, los individuos pueden ser caracterizados por su morfología: comparando especies próximas entre ellas, será posible determinar las características diferenciales constantes, por pequeñas que fuesen, que servirán como caracteres diagnósticos de cada una. De esta manera, se podrá identificar morfológicamente a los individuos, aun a aquellos muertos hace siglos.

A fines de la década de los setenta, Nadir Brum y Alicia Postiglioni, del IIBCE, que estudiaban mecanismos cromosómicos de determinación del sexo en arañas vagabundas (familia Lycosidae), nos convencieron de que sería muy fácil capturar ejemplares de la especie Lycosa thorelli, que miden unos 3cm con las patas extendidas y son frecuentes en los jardines de Montevideo (Fig. 1).
En cuanto comenzamos el trabajo, ciertos indicios nos hicieron sospechar de la existencia de más de una especie. Roberto M. Capocasale, del mismo centro de investigación, revisó los órganos genitales de las arañas capturadas pero no encontró diferencias morfológicas significativas. Intentamos cruzamientos en el laboratorio y observamos que algunos machos cortejaban intensamente a algunas hembras y poco o nada a otras; a su vez, estas hembras eran cortejadas por otros machos, que ignoraban a aquellas.
Los primeros machos mostraban siempre una aproximación cauta a la hembra, mientras agitaban patas anteriores y palpos (apéndices cefálicobucales articulados que, entre otras funciones, palpan y sujetan lo que el artrópodo come); los otros machos tenían distinto patrón de comportamiento, tan constante como el primero, en el que se alternaban quietudes absolutas con desplazamientos veloces (véase recuadro “Órganos genitales, disposiciones llavecerradura, reconocimiento y cortejo”).
Supusimos, entonces, que estábamos en presencia de dos especies, que denominamos A y B. Los machos A cortejaban con agitación y avance constantes, mientras que los B alternaban avances explosivos y pausas. Hembras A serían las que aceptaran a machos homónimos, mientras que serían hembras B las que reconociesen a los machos de esa especie. Las hembras A no aceptaban a los machos B y viceversa, lo que indicaba un reconocimiento por el comportamiento entre dos especies que coexistían en el espacio y en el tiempo. En otras palabras, constatamos el aislamiento etológico de dos especies simpátridas y sincrónicas.
Capocasale revisó cuidadosamente a los individuos, separados en lotes A y B según el comportamiento descripto, y encontró una diferencia mínima pero constante: un conjunto de pelos amarillos, microscópicos. en el primer par de patas de los machos B, que estaban ausentes en los A. Las hembras eran indistinguibles. Una de esas especies sería, entonces, la conocida, y la otra, una nueva. Como el ejemplar tipo de L. thorellidescripto en 1877 por el conde Keyserling-, que fue traido del Museo Británico al Uruguay, tenía pelos amarillos en las patas anteriores, concluimos que B era la especie identificada en el siglo pasado y A una nueva, que denominamos L. carbonelli.
En 1986, Gail Stratton y George Uetz, de la universidad de Cincinnati, advirtieron la existencia de híbridos naturales engendrados por especímenes de otras dos especies de la misma familia de arañas (Schizocosa ocreata y S. rovneri). Como ambas coexisten en algunas zonas, los híbridos indicaban que la presión selectiva no había sido lo suficientemente fuerte para evitar errores de reconocimiento, y que fallaban los mecanismos de aislamiento y los postcopulatorios. Los investigadores nombrados anularon la capacidad de reconocimiento de las hembras, anestesiándolas con dióxido de carbono (C02), y las colocaron ante machos heteroespecíficos en proceso de cortejar, los cuales realizaron cópulas más o menos normales y, por lo general, no fueron rechazados por las hembras cuando despertaron. Se generaron híbridos viables, aunque incapaces de cruzarse con individuos de las especies parentales o entre sí.
Cuando quisimos realizar experiencias similares con L. thorelli y L. carbonelli, nos encontramos ante dificultades prácticas, pues estas arañas sudamericanas son más parecidas entre ellas que lo que lo son las especies norteamericanas: el único rasgo morfológico distintivo de las primeras los pelos amarillos- es de tamaño microscópico y está restringido a los machos.
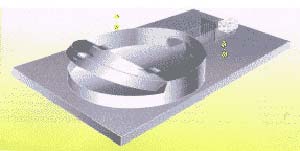
Diseñamos entonces un pequeño equipo para separar a las arañas según su comportamiento sexual y, de esta manera, distinguir las especies (Fig. 2); una vez identificados los individuos, colocamos hembras anestesiadas con C02 en el camino de machos que iniciabansu cortejo, estimulados por la feromana sexual, la substancia química producida por la hembra. Formamos cuatro grupos: dos experimentales heteroespecíficos (uno, macho A con hembra B; el otro, macho B con hembra A) y dos de control, coespecíficos (uno de cada especie).

Los machos L. carbonelli fueron más desconfiados que los L. thorelli en el momento de montarse sobre las hembras dormidas, particularmente las heteroespecíficas, y las hembras L. carbonelli se mostraron más bravías al despertar, pero, de cualquier forma, obtuvimos cópulas en todos los grupos. Las heteroespecíficas fueron atípicas, es decir, exhibieron diferencias con respecto al patrón copulatorio de los grupos de control, semejante en ambas especies y consistente en inserciones prolongadas del palpo del macho en los órganos genitales femeninos, con numerosas eyaculaciones sucesivas, fácilmente detectables por la hinchazón de un órgano elástico, la hematodocha, y la erección de las espinas de las patas del macho (Fig. 3).
Con alguna excepción, la diferencia fundamental entre los cruzamientos heteroespecíficos y los coespecíficos de los grupos de control fue la ausencia de eyaculaciones múltiples sucesivas durante una inserción. Las hembras de los grupos control generaron una prole numerosa, mientras las de los grupos heteroespecíficos no tuvieron descendencia, con la excepción de un caso, que dio origen a híbridos viables de uno y otro sexo, los cuales llegaron a la edad adulta; su existencia hizo que nos preguntáramos, con Gabriel Francescoli, de la facultad de Ciencias, si no habríamos equivocado el diagnóstico del comportamiento cuando formamos los grupos. Sin embargo, la cría misma confirmó que no hubo error. Las arañas, como los demás artrópodos, cambian la cutícula para poder crecer: L. carbonelli realizó, en promedio, ocho mudas antes de alcanzar el estado adulto; L. thorelli, diez, y los híbridos, nueve. Otras características del desarrollo, como la duración de los períodos entre las mudas, también fueron intermedias en los híbridos.
Quedaba por explicar la excepción a las cópulas atípicas. F. Pérez Miles encontró que el tamaño corporal de L. carbonelli es ligeramente mayor que el de L. thorelli. La pareja hetereoespecífíca que copuló de la manera típica se formó por azar con un macho grande de la segunda especie y una hembra pequeña de la primera. La conclusión, entonces, es que existiría una incompatibilidad entre los órganos genitales de ambas especies, la que no radicaría en la forma de estos sino en su tamaño. Por azar, una pareja superó esa restricción natural: ello permitió comprender mejor la razón del aislamiento mecánico, a la vez que confirmó la ausencia de barreras posteriores a la cópula. Quedó también confirmado que L. thorelli y L. carbonelli son especies diferentes, y que la posibilidad de híbridos naturales es prácticamente nula. De hecho, nunca hemos encontrado uno, pese a la abundancia de ambas clases de arañas en el sur del Uruguay.
Actualmente estamos analizando el comportamiento sexual de los híbridos y de los descendientes de los grupos de control. Igual que los híbridos de S. ocreata y S. rovneri, los de nuestras arañas no logran copular entre ellos ni con integrantes de las especies parentales. Comparando el comportamiento sexual de los diferentes grupos, encontramos indicios de que estamos entrando en un terreno promisorio en el estudio de la herencia de las formas de cortejo. Pero esto, como hubiese dicho Rudyard Kipling, es otro cuento…




