




La mulita, al igual que otros armadillos, gesta crías genéticamente idénticas que provienen de un único óvulo fecundado. Este fenómeno se conoce como poliembrionía, que también da lugar a los gemelos univitelinos humanos.
En 1915, Miguel Fernández (1882-1950), profesor del Museo de La Plata, publicó un exhaustivo trabajo de investigación sobre la reproducción en la mulita (Dasypus hybridus) en el cual presentó evidencia acabada de un extraordinario fenómeno al que se había referido en una publicación de 1909: la gestación por un mamífero de múltiples crías genéticamente idénticas, llamado poliembrionía. Informó que dicha mulita gestaba durante su preñez siete, ocho, once o doce crías idénticas. Ese fenómeno ya había sido señalado por zoólogos que observaron, en otras especies, camadas numerosas de crías del mismo sexo, pero ninguno lo había estudiado y nada se sabía acerca de sus mecanismos.

El artículo de Fernández atrajo inmediatamente la atención mundial de zoólogos, embriólogos y biólogos. Lo publicó en alemán, como era costumbre en la época, con un amplio resumen en castellano, en el Morphologisches Jahrbuch. Incluía un capítulo escrito por su esposa, Katy Marcinowski, sobre el desarrollo del sistema nervioso de ese particular armadillo.
Miguel Fernández, científico
Miguel Fernández nació el 25 de septiembre de 1882 en Essen, en la región del Ruhr, de padre argentino y madre alemana. La familia se estableció poco después en el Uruguay. Miguel hizo sus estudios primarios y comenzó los estudios secundarios en Montevideo, que continuó en Suiza. Completó su bachillerato en el politécnico de Zurich y luego, en la Universidad de Zurich, se doctoró summa cum laude en zoología, bajo la guía de Arnold Lang. Continuó su formación científica en la Universidad de Heidelberg.

Cuando el paleontólogo Santiago Roth, del Museo de La Plata, consultó a Lang acerca de un zoólogo para la institución, este recomendó entusiastamente al joven Miguel Fernández, a quien consideró uno de sus mejores discípulos. En 1906, el director del museo, Samuel Lafone Quevedo, invitó a Fernández, entonces de veintitrés años, a asumir el cargo de profesor de zoología y jefe de la sección correspondiente del museo. Ya había publicado en una importante revista alemana un trabajo sobre la histología de los tunicados y había adquirido gran versación en los métodos experimentales más modernos de la época.
Luego de su designación como profesor de zoología de la Universidad Nacional de La Plata, comenzó sus estudios de la fauna argentina de mamíferos; centró su atención en los armadillos y especialmente en la mulita. En 1909 publicó el trabajo mencionado en el texto sobre algunos aspectos de la embriología de ese animal. En julio de 1910, comunicó sus observaciones con más detalles al Congreso Científico Internacional Americano, organizado por la Sociedad Científica en Buenos Aires como parte de los actos del Centenario de la Revolución de Mayo.
En 1915 salió su trabajo fundamental sobre la poliembrionía, que ocupó la totalidad del volumen 21 de la Revista del Museo de La Plata. Voluminoso, exhaustivo y finamente detallado, con 169 figuras y láminas explicativas, el trabajo constituyó uno de los más importantes pilares de la biología del desarrollo en el país.
Durante todo el tiempo que permaneció en el Museo de La Plata, Fernández trabajó con exclusividad en docencia e investigación. En 1926 renunció a sus cátedras de Zoología y Anatomía Comparada en el Museo de La Plata y pasó a revistar con ese mismo rango en la Universidad Nacional de Córdoba. Allí publicó su trabajo sobre histología y embriología del exoesqueleto del peludo (Chaetophractus villosus) en las Actas de la Academia Nacional de Ciencias (1931). Por otra parte, difundió los más recientes aportes a la genética (‘El transformismo y las proyecciones de la genética sobre el mismo’, Cursos y Conferencias, Colegio Libre de Estudios Superiores, 1934). Fue seguramente uno de los primeros que se dedicaron a la genética en la Argentina, y su interés se prolongó en la trayectoria de sus discípulos, en especial Francisco A Sáez. Ese interés y sus propias ideas sobre genética impulsaron a Fernández a escribir un libro sobre el tema: Genética general humana y elementos de biometría, junto con su esposa Katy Marcinowski. Después de su muerte en 1950, fue publicado por esta (LEDS, Córdoba, 1956).
Fernández demostró que en esa clase de parición todas las crías provienen de un único óvulo fecundado, conocido como cigota o célula huevo. Esto explica que las crías sean idénticas, ya que poseen el mismo genoma y, lógicamente, también explica que las crías de una camada sean siempre del mismo sexo. Fernández descubrió que la poliembrionía no se origina en una separación temprana de células embrionarias para formar embriones distintos, sino en un estadio posterior, cuando ya están diferenciadas las primeras estructuras embrionarias, en el estadio que se conoce como embrioblasto. En ese momento del desarrollo se forman el disco embrionario, que da lugar al feto, y el trofoblasto, que da lugar al tejido extraembrionario, como la placenta. Esta demostración requirió de un laborioso examen de secciones seriadas y de reconstrucción espacial de embriones de distintas edades tempranas, lo cual demostró en el científico una excepcional calidad técnica y gran imaginación creativa en la interpretación de sus datos.
Resulta extraordinario que la capacidad de generar un embrión completo, que en los mamíferos normalmente se reserva a las primeras dos o cuatro células, esté presente en un estadio del desarrollo mucho más avanzado, como lo está en las células de cada disco embrionario de la mulita.
La poliembrionía
Si se observa el abdomen abierto de una mulita preñada al principio de la gestación, se puede distinguir una placenta única a la cual están conectados por cordones umbilicales siete, ocho, once o doce embriones, como se muestra en la figura 1. Todos los embriones son idénticos y este hecho ocurre tanto en animales silvestres como en cautiverio, lo que descarta causas ambientales.
Histología del embrión de la mulita
Como parte de sus estudios, Fernández documentó sus observaciones con microfotografías y fotografías convencionales de los fetos y la placenta. Logro compilar un relato completo del desarrollo de la mulita con un grado de detalle superior al de los textos de embriología en uso, dado que trabajó con cortes seriados y reconstrucciones espaciales hechas de cartón.
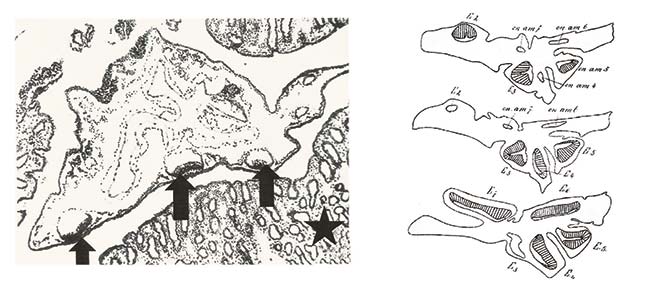 A la izquierda, microfotografía de un embrión no implantado tomada con un aumento de 50 diámetros en el microscopio. Las flechas indican tres discos embrionarios con sus cavidades amnióticas; la estrella señala la mucosa uterina. A la derecha, cortes de tres vesículas embrionarias de mulita con los discos embrionarios resaltados con rayado. Fernández publicó ambas ilustraciones en el volumen 21 de la Revista del Museo de La Plata, 1915.
A la izquierda, microfotografía de un embrión no implantado tomada con un aumento de 50 diámetros en el microscopio. Las flechas indican tres discos embrionarios con sus cavidades amnióticas; la estrella señala la mucosa uterina. A la derecha, cortes de tres vesículas embrionarias de mulita con los discos embrionarios resaltados con rayado. Fernández publicó ambas ilustraciones en el volumen 21 de la Revista del Museo de La Plata, 1915.
El fenómeno es parecido al que sucede en los humanos cuando nacen gemelos univitelinos o monocigóticos, lo que tiene una frecuencia promedio de uno en cada 290 nacimientos. Se suponía erróneamente que este tipo de gemelos humanos se formaba debido a la separación de dos células embrionarias muy tempranas, pero gracias a los trabajos de Fernández se conoce que es un evento posterior.
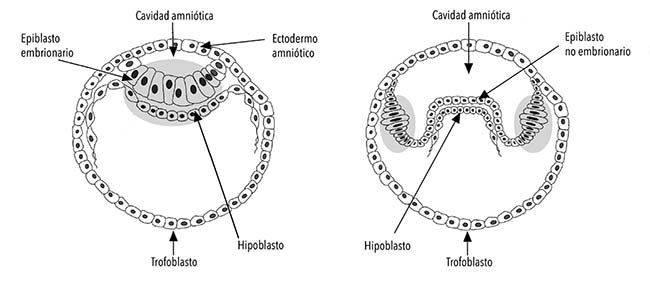 A la izquierda, el desarrollo temprano convencional de un embrión de mamífero; a la derecha, el desarrollo temprano en la mulita. Las manchas grises son los discos embrionarios que en cada caso dan lugar a los embriones. La capa celular superior, llamada epiblasto, forma el piso de la cavidad amniótica; la capa inferior, hipoblasto o endoblasto, forma el techo del saco vitelino. En la mulita, se indica la formación de pequeñas bolsas alargadas en forma de dedo de guante, una para cada embrión, que comparten la cavidad amniótica con su líquido (solo se representa dos bolsitas).
A la izquierda, el desarrollo temprano convencional de un embrión de mamífero; a la derecha, el desarrollo temprano en la mulita. Las manchas grises son los discos embrionarios que en cada caso dan lugar a los embriones. La capa celular superior, llamada epiblasto, forma el piso de la cavidad amniótica; la capa inferior, hipoblasto o endoblasto, forma el techo del saco vitelino. En la mulita, se indica la formación de pequeñas bolsas alargadas en forma de dedo de guante, una para cada embrión, que comparten la cavidad amniótica con su líquido (solo se representa dos bolsitas).
La descripción de las etapas sucesivas que originan la poliembrionía en la mulita se enriquecieron por el debate con los zoólogos estadounidenses Horatio H Newman (1875-1957), de la Universidad de Chicago, y John Thomas Patterson (1878-1960), de la Universidad de Texas, quienes, estimulados por el primer trabajo de Fernández, estudiaron otra especie de armadillo, la mulita grande (Dasypus novemcinctus) presente en Texas, que exhibe la particularidad de tener camadas de cuatro crías idénticas. Sostuvieron que cada uno de los embriones deriva de las cuatro células luego de la segunda división del cigoto.
Fernández determinó que las primeras etapas del desarrollo embrionario de la mulita son similares a las de los demás mamíferos, hasta la formación del embrioblasto o vesícula embrionaria, como se muestra en la figura 2. En este estadio, se diferencian el trofoblasto (la capa periférica de células), una cavidad inicial y una masa celular interna en la cual se produce otra cavidad más pequeña, la del amnios primario, y se forma la estructura, conocida como el disco embrionario, cuya capa superior de células se llama epiblasto. Ese disco da lugar al embrión.
En los armadillos, en ese estadio del desarrollo, cuando hay centenares de células derivadas del huevo o cigota, ocurren los fenómenos que resultan en la poliembrionía, y que diferencian a esos animales de los demás mamíferos estudiados. En la mulita, las células que forman el epiblasto proliferan y constituyen en su periferia un número de pequeñas bolsas o depresiones con forma de dedo de guante. La cantidad de bolsitas corresponde al futuro número de embriones y, finalmente, de crías. Es decir, en un momento en que hay varios centenares de células embrionarias, las del epiblasto original dan lugar a varios nuevos discos embrionarios en las bolsitas creadas, y cada bolsita forma un embrión. Estas observaciones implican que todas esas células epiblásticas tienen la capacidad de inducir la formación de un individuo completo, es decir, son pluripotenciales.
Las observaciones de Fernández estuvieron fundamentadas en micrografías y cortes histológicos, algunos de los cuales se muestran en el recuadro ‘Histología del embrión de la mulita’.
Repercusiones del descubrimiento
La poliembrionía es un fenómeno natural que revela la capacidad de ciertas células embrionarias de organizar un organismo completo. En ese aspecto se relaciona con la potencialidad de las células madre embrionarias, un tema que es objeto de numerosas investigaciones actuales dado que hay dos componentes en el embrión: el embrión propiamente dicho y el tejido extraembrionario, que finalmente forma la placenta. La diferenciación de linajes celulares para formar el feto y los tejidos extraembrionarios es el primer cambio que ocurre en un embrión. En los últimos años esto se ha estudiado usando exclusivamente unos pocos mamíferos de laboratorio, en especial ratones. En esos experimentos, se observa una pérdida progresiva de la potencialidad desde el huevo hasta la quinta división sucesiva de las células del embrión. La separación de células con la potencialidad de generar un embrión completo se ha demostrado experimentalmente en algunos mamíferos, pero solo los armadillos y los seres humanos muestran la ocurrencia regular de los gemelos univitelinos.
Varios autores han señalado las similitudes entre la poliembrionía en armadillos y en humanos. El estudio de embriones humanos está restringido por cuestiones éticas y por regulaciones legales, por lo que el mecanismo que lleva a la formación de los gemelos univitelinos no está comprobado fehacientemente.
En cambio, los armadillos ofrecen un material excepcional para investigar las variaciones de la potencialidad de las células embrionarias. Como lo estableció Fernández en 1915, la partición de las células del epiblasto origina la poliembrionía y, por consiguiente, esas células tendrían la actualmente llamada pluripotencialidad primitiva o potencialidad para forman el cuerpo completo de un embrión que compartirá el tejido extraembrionario, es decir, la placenta, con sus gemelos.
Más allá de la poliembrionía
Las mulitas son animales de indudable interés científico. También lo tiene el estudio tanto de su desarrollo gonadal (de sus testículos, ovarios y las correspondientes vías) como de su placenta. En los últimos dos decenios se han realizado investigaciones sobre esos temas y se está integrando rápidamente el estudio de la biología de la reproducción de los armadillos mencionados en este artículo.
Los autores agradecen a Irina Podgorny, Susana García y Jorge Affani su interés y sus aportes a este artículo.
Alberto J Solari
[email protected]




