




Histología de osteodermos
El registro fósil de vertebrados está representado principalmente por la presencia de piezas óseas. Estas incluyen huesos del esqueleto, tendones osificados, dientes y osteodermos. Los osteodermos, también denominados oscículos o placas, son estructuras óseas localizadas en la piel o tegumento. La piel está constituida por dos capas: una superficial (la epidermis) y otra más profunda, la dermis, donde se encuentran los osteodermos. Estas estructuras aparecen en vertebrados fósiles y vivientes, y presentan diferentes formas, extensiones, ornamentaciones, tamaños y funciones. Este artículo se enfoca en las implicancias fisiológicas y ecológicas que surgen de los osteodermos a partir de estudios comparativos de la microestructura interna de sus tejidos (histología) en grupos extinguidos y actuales de tetrápodos (vertebrados de cuatro extremidades). Si bien suele denominarse ‘armadura’ o ‘coraza’ a la cubierta de osteodermos, esta denominación es técnicamente incorrecta, ya que denota una función de protección implícita a priori. No obstante, las funciones pueden ser más amplias que únicamente como estructura defensiva. La técnica convencional para examinar la histología de elementos óseos es mediante secciones delgadas. Estas consisten en porciones de hueso cortadas y pulidas hasta llegar a un espesor de unas decenas de micrones (1 micrón = 0,001mm) para que puedan ser observadas bajo lupa binocular o microscopio con luz transmitida.
¿DE QUÉ SE TRATA?
Sobre los osteodermos, estructuras que se encuentran en la piel de animales tetrápodos fósiles y vivientes. Su función y las inferencias que se obtienen del estudio en fósiles.
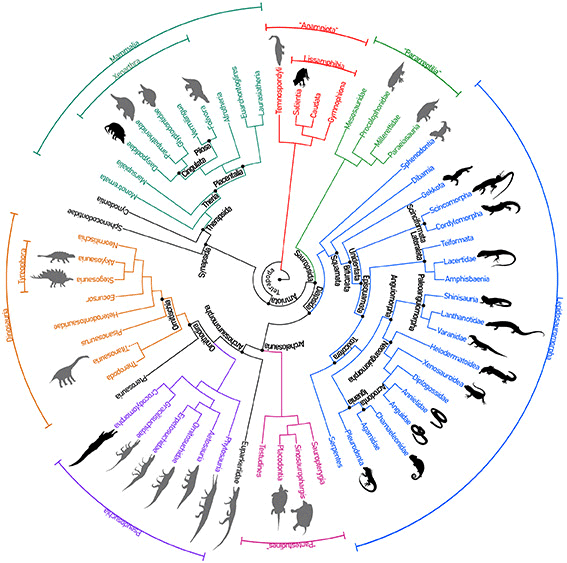
Los osteodermos aparecieron tempranamente en la evolución de los tetrápodos, durante el Devónico Superior, hace más de 385 millones de años (Ma). Entre los representantes más primitivos se encuentran los temnospóndilos (extinguidos) y los actuales anfibios (ranas y sapos entre otros), de hábitos anfibios o acuáticos. Posteriormente, en el período Carbonífero (hace unos 340Ma), aparecieron los amniotas, tetrápodos caracterizados por poseer huevos con cuatro envolturas protectoras, rasgo que les permitió independizarse del agua y desarrollarse en ambientes terrestres. Entre los amniotas vivientes pueden identificarse, en el linaje de reptiles, varios grupos de lepidosauromorfos portadores de osteodermos (camaleones, geckos, cordílidos). Otro grupo de importancia con armadura dérmica son los cocodrilos y caimanes. Finalmente, entre los mamíferos, se incluyen los xenartros, cuyo grupo acorazado por excelencia son los armadillos. Si se abarcan grupos fósiles, el panorama se hace más extenso y complejo. Por ejemplo, entre los xenartros fósiles pueden incluirse a los gliptodontes y pampatéridos, grandes formas con una coraza en forma de cúpula. Los milodontes fueron otro grupo de xenatros (con osteodermos que formaban una escasa armadura) relacionados con los perezosos modernos (folívoros). Por otra parte, en el linaje que incluye a los cocodrilos (pseudosuquios), además de representantes fósiles de estos, existieron varios grupos, tales como aetosaurios, fitosaurios y ornitosuquios con osteodermos de variada forma y extensión. En el linaje hermano de los pseudosuquios, en los ornitodiros, algunos dinosaurios como anquilosaurios y estegosaurios (tireóforos) y saurópodos especializados (titanosaurios) poseían una armadura de osteodermos, en mayor o menor medida. Entre algunos grupos fósiles escasamente conocidos con una armadura dérmica puede mencionarse a los pareiasaurios, procolofónidos y placodontos, este último vinculado a las tortugas modernas. Sin embargo, cabe destacar que las placas del caparazón de las tortugas no pueden considerarse osteodermos verdaderos porque tienen un origen embrionario diferente.

¿Cómo se forman? ¿Cómo están constituidos?
Los osteodermos están constituidos principalmente por tejido óseo, al igual que los huesos del esqueleto, así que en general se emplean los mismos términos anatómicos para describirlos (ver Cerda, 2018, en las lecturas sugeridas, donde se recopilaron los aspectos más importantes que caracterizan al tejido óseo). Con respecto a su formación, principalmente se ven involucrados dos mecanismos: osificación intramembranosa y osificación metaplásica. En la primera, el hueso se forma a partir de la condensación y multiplicación de células especializadas desde un tejido conectivo primitivo. La metaplasia consiste en la transformación directa de un tejido blando en tejido óseo sin la intervención de células especializadas o molde previo de cartílago. Esta última es reconocida en fósiles por la presencia de haces densamente empaquetados de fibras colágenas mineralizadas orientadas en dos o tres direcciones y denominadas comúnmente ‘fibras estructurales’. La ocurrencia de un tipo u otro de osificación parecía ser mutuamente excluyente. Por ejemplo, se ha inferido que los osteodermos de la mayoría de los armadillos se desarrollan por osificación intramembranosa y los de los lagartos, por metaplasia. Sin embargo, en estudios recientes efectuados en cocodrilos (Alligator mississippiensis) se reconoció la intervención de ambos tipos de osificación desde etapas embrionarias. Asimismo, en grupos fósiles se propone un modo mixto de osificación en los osteodermos de algunos arcosauromorfos. Finalmente, se ha inferido un tercer modo de osificación en fósiles, el cual se da a partir de un molde de cartílago, registrado en los osteodermos de placodontos (reptiles marinos).
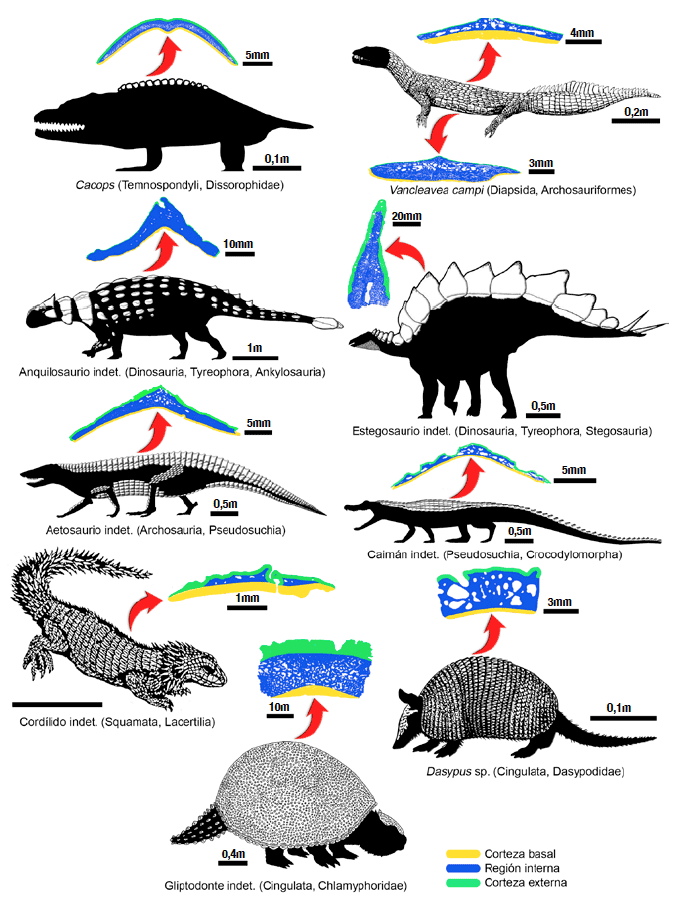
Estructuralmente, la mayoría de los osteodermos se disponen en tres capas de tejidos, que pueden ser caracterizados según su porosidad relativa en hueso compacto o hueso esponjoso. El primero está caracterizado por una baja porosidad: el tejido óseo es más abundante con relación a la cantidad de espacios, canales y cavidades (poros). En el hueso esponjoso, la relación es inversa: los poros ocupan mayor espacio que el tejido óseo. Considerando la estructura interna de los osteodermos, se suelen dividir en una corteza basal (o profunda, orientada hacia el interior del organismo) de hueso compacto, una intermedia de hueso esponjoso (región medular) y otra corteza externa (o superficial, orientada hacia la epidermis) de hueso compacto. En algunas especies de lagartos (esquincos, geckos), se presenta además una cuarta capa, sobre la corteza externa, compuesta por un tejido denso particular denominado osteodermina, la cual posee aspecto vítreo, similar al esmalte de los dientes.
¿Cuál es su función?
La respuesta más aparente respecto de su función es que sirven como estructura defensiva. Los dinosaurios del grupo de los anquilosaurios ilustran perfectamente esta cuestión. Estos fueron dinosaurios cuadrúpedos que vivieron del Jurásico al Cretácico (167-66Ma). Podían alcanzar hasta 8 metros de largo, presentaban miembros cortos y robustos y una cabeza pequeña, poseían una extensa cobertura de hileras de osteodermos de formas variadas. Algunos coronaban el extremo de su cola con un prominente mazo óseo formado, precisamente, por osteodermos.
Si bien la utilidad de la armadura como elemento defensivo parece ser cierta para tetrápodos fuertemente acorazados, la cuestión sobre la funcionalidad es más compleja. En primer lugar, hay especies, actuales y extinguidas, cuya escasa cantidad y/o densidad de osteodermos hace imposible considerarlos como mecanismo de defensa. En algunos anuros, por ejemplo, la rana mono (Phyllomedusa vaillanti) de Perú, se observa una hilera de diminutos osteodermos en las porciones laterales de su cuerpo. Entre los lagartos, por mencionar a una especie, el camaleón de hoja antsingy (Brokesia perarmata) de Madagascar presenta una hilera dorsal continua de osteodermos con pronunciadas proyecciones y dos hileras costales de osteodermos dispersos.
Del mismo modo, la densidad y distribución de osteodermos en algunos grupos fósiles de tetrápodos contradicen claramente una función defensiva para estas estructuras. Entre ellos, se pueden mencionar a los temnospóndilos disorófidos del Pérmico (300-250Ma) de América del Norte. De pequeño tamaño (hasta 40cm de largo) y morfología similar a las salamandras, poseían dos hileras de osteodermos dorsales. Algunos dinosaurios, como los estegosaurios, que tuvieron una diversificación global desde el Jurásico al Cretácico (170-125Ma), también pueden incluirse entre formas con una armadura dispar. Estos eran herbívoros cuadrúpedos de gran tamaño (hasta 9 metros de largo), con miembros delanteros cortos, miembros traseros largos y una cabeza alargada y pequeña. Sus características más distintivas son las dos hileras dorsales de grandes placas óseas generalmente subtriangulares. Algunos portaban largas púas sobre el extremo de la cola.
Dada la extensión de la armadura dérmica en los taxones antes mencionados, su rol como elemento defensivo es en muchos casos muy poco evidente. Si bien en la rana mono no se conoce otra función más allá de la defensa pasiva, en el camaleón de hoja antsingy se cree que los osteodermos sirven para el camuflaje. Además, la compleja estructura de la armadura de este último sería útil para impedir ser tragados por depredadores. Entre los grupos fósiles, los osteodermos en los disorófidos pudieron servir como estabilizadores de la delicada columna durante la transición hacia el medio terrestre. En el caso de los estegosaurios, mientras que la presencia de placas ha sido relacionada a un posible dimorfismo sexual o como elementos termorreguladores, las púas caudales se consideraron como elementos utilizados para la defensa activa contra depredadores.
Asimismo, en formas que presentan una armadura extensa, esta puede tener otras funciones además de la protectora. En este sentido, en los cocodrilos los osteodermos sirven como depósito de iones de calcio para su movilización en la ovogénesis (formación de los huevos), para evitar la acidificación del organismo durante las inmersiones (por la falta de respiración), para la transferencia de calor durante el asolamiento y como puntos de anclaje de la musculatura. En los mamíferos, la armadura de los armadillos está asociada a diferentes glándulas, que podrían estar vinculadas a adaptaciones medioambientales. Los aetosaurios, reptiles cuadrúpedos y herbívoros del Triásico (250-200Ma), son de particular interés por sus tipos de osteodermos. Estaban fuertemente acorazados con varias hileras de osteodermos dorsales, laterales e incluso ventrales, con una pronunciada ornamentación consistente en crestas y surcos. Se ha propuesto que el grado de ornamentación y el tamaño de sus osteodermos eran indicativos de dimorfismo sexual.
¿Qué información aportan?
Reconstrucciones anatómicas. La forma en que se disponen los osteodermos y su microestructura permitieron inferir en algunas especies la distribución de tejidos blandos (músculos, ligamentos). Desde la histología, la presencia de tejidos blandos en fósiles es deducida a partir de las fibras de Sharpey (haces delgados y estrechamente agrupados de fibras mineralizadas de colágeno). En croniosuquios, tetrápodos basales del Pérmico de Rusia con una armadura prominente, la presencia de estas fibras permitió deducir que los osteodermos están conectados a las vértebras a través de la musculatura, tal como ocurre en muchos grupos de cocodrilos actuales.

Clasificación. Algunos osteodermos presentan particularidades exclusivas para determinados grupos. Los dasipódidos, cingulados que incluyen a los armadillos actuales, reflejan estas implicancias. Al tratarse de animales vivientes, pueden observarse características relativas tanto del tejido óseo como del blando. Los dasipódidos se dividen en tres subfamilias: dasipodinos (mulitas), eufractinos (peludos) y tolipeutinos (tatú carreta). El grado de densidad o compactación de los osteodermos (cavidades), la distribución de las fibras de Sharpey, la ocurrencia y abundancia de glándulas asociadas (sudoríparas, secretoras de grasa y productoras de pelos) son aspectos que permiten diferenciar al menos a las primeras dos subfamilias. Del mismo modo, se han empleado osteodermos de anquilosaurios para desarrollar esta clase de estudios. Se observó que la extensión de la corteza basal y la orientación de las fibras estructurales parecen ser características constantes para las tres familias de anquilosaurios: polacántidos, anquilosáuridos y nodosáuridos. Esta herramienta resulta útil en grupos fósiles, ya que las fuentes de información son escasas. Además, por lo general, en formas acorazadas, los osteodermos se hallan mejor preservados (respecto de la cantidad) que los huesos del resto del esqueleto.
Edad. El tejido óseo posee estructuras específicas que surgen de la variación en la tasa de crecimiento, tanto a nivel morfológico como histológico. Estas son llamadas marcas de crecimiento. Dado el carácter anual de su formación, resultan de gran utilidad para determinar la edad de los individuos. Así, pueden mencionarse las líneas de detención del crecimiento (LDC), las cuales indican una pausa en el crecimiento. Otras marcas de crecimiento son los annuli y las zonas, las cuales reflejan períodos con tasas bajas y altas de crecimiento respectivamente. Un grupo fósil que ha demostrado cierta fiabilidad como indicador de edad (mínima) en sus osteodermos son los erpetosuquios, cuadrúpedos carnívoros de postura erguida del Triásico.
Sin embargo, son necesarios ciertos recaudos para la estimación. Por ejemplo, como se mencionó anteriormente, durante su etapa reproductiva las hembras de cocodrilos reabsorben una parte de sus osteodermos (remodelación ósea) para movilizar el calcio de estos durante la ovogénesis. En este proceso también se remueven posibles marcas de crecimiento. Además, los cocodrilos comienzan a depositar marcas de crecimiento un año después de su eclosión. Por estas cuestiones, los osteodermos que son más útiles como indicadores de edad son aquellos con reducida o nula remodelación. Finalmente, combinando esta información con otros parámetros anatómicos (largo o ancho de los osteodermos, por ejemplo) también se han intentado reconstruir curvas de crecimiento (gráficos que ilustran cómo crece un individuo a lo largo del tiempo), como es el caso de algunos aetosaurios y cocodrilos.
Paleoecología. Estudios morfológicos y microestructurales de la armadura dérmica permiten conocer aspectos ligados a la relación de los individuos con su ambiente y con otros organismos. Se ha observado que los huesos de especies actuales y de hábitos acuáticos o anfibios son típicamente más densos y macizos que aquellos de animales terrestres. Esta característica se ha considerado como una adaptación que facilita la flotabilidad (a modo de lastre). Estas relaciones causales entre la microestructura ósea de los osteodermos y el hábito han sido empleadas para inferir este último en especies fósiles. Así, por ejemplo, el arcosauromorfo Vanclevea campi, del Triásico Superior (237-201Ma) de América del Norte (de morfología superficial similar a la de una iguana marina), cuyo cuerpo estaba completamente rodeado por cinco diferentes formas de osteodermos, poseía oscículos fuertemente macizos. Esto sugiere que la coraza pudo haber funcionado como ‘lastre’, facilitando la locomoción acuática del organismo. No obstante, estudios recientes valoran esta deducción un tanto reduccionista, sobre todo para formas fósiles con estilos de vida inciertos. Puede derivarse, sin embargo, que el grado de compactación ósea de los osteodermos es un dato complementario a estudios morfológicos y microanatómicos para apelar a un hábito determinado.
Consideraciones finales
Los estudios de osteodermos se han incrementado en los últimos años, principalmente en grupos fósiles. Nuevos aportes efectuados por paleontólogos intentan examinar estos tópicos con un mayor número de muestras para robustecer los resultados obtenidos. Asimismo, se han comenzado a analizar los resultados mediante métodos estadísticos, transformando las variables cualitativas a cuantitativas, para apuntar a procedimientos más precisos. Al mismo tiempo, se emplean otras técnicas para explorar los osteodermos a nivel ultraestructural (por ejemplo, análisis con microscopio electrónico de barrido) y extraer otras características de importancia. Estos datos, en conjunto con otras fuentes de información (morfológicas), permitirán ajustar mejor el tipo de inferencias que pueden hacerse en especies extinguidas a partir del estudio de sus osteodermos.
Agradezco a Ignacio Cerda y Julia Desojo por su asesoramiento en la preparación de este artículo.
LECTURAS SUGERIDAS
CERDA IA, 2018, ‘Mirando gigantes con lupa’, Ciencia Hoy, 27 (159): 13-19.
HALL BK, 2015, ‘Variation outside the norm: Neomorphs and atavisms’, en Hall HK (ed.), Bones and Cartilage Developmental and Evolutionary Skeletal Biology, Elsevier, La Haya, pp. 693-707.
PADIAN K & LAMM ET, 2013, Bone Histology of Fossil Tetrapods, University of California Press.

Licenciado en paleontología, UNRN.
Becario doctoral de Conicet en el Instituto de Investigación en Paleobiología y Geología (IIPG), UNRN.
Docente auxiliar, UNRN.



