




La estrucutura fractal de algunos componentes de los sistemas vivos permite explicar por qué ciertos procesos biológicos se comportan, dinámicamente, en forma diferente de otros.
Los procesos que tienen lugar en los organismos vivos abarcan amplias escalas de espacio y de tiempo. La relación entre estas dos variables es una medida de la velocidad con la que ocurren. Sin embargo, la relación no es uniforme y, en determinado momento, se manifiestan fenómenos de coherencia que encuentran una explicación al estudiar la geometría de las estructuras involucradas.
Existen muchos fenómenos muy comunes en el mundo natural que no se pueden describir utilizando las herramientas que la física emplea para explicar el movimiento del péndulo, o la termodinámica tradicional para estudiar los sistemas en equilibrio. Entre ellos pueden mencionarse los latidos del corazón; las distribuciones ‘regionalizadas’ de los depredadores en los ecosistemas; el comportamiento caótico del tiempo atmosférico; la descripción, utilizando dimensiones fraccionarias, de objetos cuyas formas no cambian al cambiar la escala con la que se los mira -como las irregularidades de una costa-, o la misma estructura de las alvéolos pulmonares. Todas ellas comparten la propiedad de exhibir un cierto comportamiento coherente, o sea que las distintas partes del conjunto son similares en escalas distintas de longitud ( invariancia de escala espacial ) o desplie- gan la misma actividad simultáneamente ( invariancia de escala temponaI ), de manera análoga a como, en el latido, las diferentes fibras del músculo cardiaco se contraen al mismo tiempo. Por otro lado, estos fenómenos sólo se manifiestan bajo ciertas circunstancias muy particulares, tan excepcionales coma las que hacen posible la vida en nuestro planeta.
En el presente articulo vamos a considerar los fundamentos teóricos y experimentales de la organización dinámica -espacial y temporal- de los organismos vivos, y lo ilustraremos con el ejemplo específico de la organización del citoplasma y de los procesos que ocurren en él.
Niveles de organización y mecanismos de coherencia de la función celular
El orden característico de los seres vivos, en el espacio y en el tiempo, surge de la dinámica misma de la organización de los procesos, de los cuales podemos encontrar ejemplos en nuestra experiencia cotidiana: el movimiento del aire que fluye en los pulmones, la renovación permanente de las células hepáticas, la circulación de la sangre, impulsada por el ritmo del corazón, el movimiento de iones que estimulan los impulsos eléctricos en las células nerviosas, el ritmo menstrual femenino, entre otros. De estos movimientos o flujos depende la continuidad de nuestra vida; lo mismo ocurre en cada nivel de organización de las células en todos los organismos.
Uno de los antecedentes conceptuales inmediatos de la organización dinámica (Figs. 1 y 2) es el reconocimiento de varios niveles de organización en el funcionamiento de los organismos vivientes. A estos niveles podemos jerarquizarlos de acuerdo con un criterio de complejidad creciente. Por ejemplo, en un nivel inicial podemos ubicar a las moléculas, como las proteínas; luego a las ‘asociaciones’ moleculares, que forman los complejos de proteínas; luego, a las estructuras subcelulares, que constituyen las mitocondrias; después, las células, por ejemplo las epiteliales, los tejidos, en este caso la epidermis; los órganos, aquí la piel; y siguen los sistemas, los organismos, y, por último, las poblaciones.
Nos planteamos efectuar una caracterización cuantitativa de estos niveles de organización, para lo cual nos hemos concentrado en los procesos biológicos que tienen lugar en cada uno de ellos, como -por ejemplo- los existentes en la regulación, por las proteínas, de la velocidad de las reacciones biológicas que tienen lugar en el primero de los niveles, el molecular. Realizamos así una caracterización del tiempo y del espacio en el que ocurren los distintos procesos biológicos correspondientes a cada nivel (véase recuadro ‘Niveles de organización biológica’).
El funcionamiento de distintos procesos celulares opera simultáneamente en varios niveles de organización, caracterizados por distintas escalas de espacio y de tiempo, como se explica en el citado recuadro. Aun cuando dichos niveles pueden ser muy dispares entre sí en cuanto a las dimensiones de espacio y tiempo en las que operan, están profundamente relacionados y tienen entre ellos una continuidad física que describiremos enseguida (véase recuadro ‘La autoorganización espacio-temporal’ y la Fig. 3).

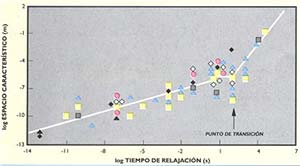
En el gráfico de la figura 1 se han representado los tiempos característicos de los procesos que ocurren en distintos niveles de organización, así como los espacios en los que se llevan a cabo (véase recuadro ‘Niveles…’). Si se traza una recta que pase por la mayoría de los puntos se observa un quiebre en la pendiente de esta a la altura de los minutos (en la escala temporal) y de los micrómetros (en la espacial). Este cambio de pendiente, que hemos llamado punto de transición, ha sido interpretado por quienes lo describieron por primera vez como aquel a partir del cual aparecen comportamientos coherentes a nivel macroscópico, es decir, observables a simple vista o con poco aumento. Esta coherencia requiere que los movimientos o actividades de muchas partes, en procesos aparentemente independientes, se coordinen entre sí para dar un único comportamiento organizado. Este comportamiento coherente se contrapone con el comportamiento al azar, en el que las partes actúan independientemente y de forma distinta, sin ningún orden ni coordinación entre ellas.
Un hecho para destacar es que este punto de transición corresponde a las dimensiones de espacio y de tiempo en que las estructuras del citoesqueleto (microtúbulos y microfilamentos) se arman y se desarman por montaje o desmontaje de las proteínas que lo forman (véase ‘La arquitectura de las neuronas’, Ciencia Hoy 3:34, 1989). Esta coincidencia de coordenadas de espacio y de tiempo nos ha llevado a postular que los procesos de montaje y desmontaje del citoesqueleto están involucrados en la coordinación de actividades en la organización celular. Como consecuencia, las reacciones bioquímicas serían sensibles a esos procesos de armado y desarmado, los cuales, por tener lugar en escalas del orden de los micrómetros, permitirían coordinar y sincronizar procesos bioquímicos en sitios muy distantes en el interior celular. A partir de este punto, nos centraremos en la organización del citoplasma, donde ocurren estos procesos, e ilustraremos con un ejemplo concreto el concepto de organización dinámica (véase recuadro ‘Otros modelos de arquitectura y función citoplasmática’).
Redes moleculares fractales. El citoplasma como un fractal de percolación
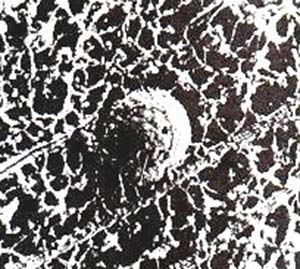
Actualmente, se cree que las reacciones químicas del metabolismo celular no ocurren en un medio acuoso homogéneo, sino en uno estructurado, como es el citoplasma (Fig. 4). La estructura que probablemente organiza las actividades bioquímicas, al brindar un soporte para las proteínas (enzimas) que aceleran las reacciones, es el esqueleto celular; en él se pueden distinguir dos niveles de organización: el sistema microtrabecular y el llamado citoesqueleto. Ambos están formados por proteínas que se disponen en forma de filamentos y entrecruzadas (Fig. 4).

Una primera observación relevante es que el análisis de la geometría del citoesqueleto y del sistema microtrabecular revela la naturaleza fractal de su tramado (véase recuadro ‘Objetos fractales’). Consideremos ahora qué tipo de fractal podría ser el citoplasma celular. Basándonos en la determinación de la dimensión fractal, D, a partir de fotografías de microscopia electrónica, tanto la malla microtubular (uno de los elementos del citoesqueleto) como el sistema microtrabecular arrojan valores de D=1,89. En la figura 4, por otro lado, se muestra una foto, obtenida por microscopia electrónica, del citoplasma de una célula nerviosa; en ella, se distinguen los filamentos de las proteínas que componen el esqueleto celular, conectados entre sí por proteínas asociadas. Analizando convenientemente estas fotos, puede deducirse la dimensión fractal de estas estructuras, de las que se obtiene D=1,9072, o sea que es muy similar a la anterior y ambas muy próximas a la dimensión que corresponde a lo que se llama agregado de percolación. Los fractales de percolación se denominan así por analogía con el comportamiento del café al ser filtrado (en inglés, percolation). El término percolación hace referencia a un proceso de difusión sólo a través de ciertos sitios en el espacio (los poros del filtro), dejando la imagen de un conjunto irregular de esos sitios mojados por el café (el agregado o cluster de percolación).

El valor determinado de D corresponde a imágenes bidimensionales (2D) de un espacio tridimensional (3D). Dado que la red macromolecular del citoplasma es 3D, es de esperar que la dimensión (D) medida sea mayor que dos y menor que tres, o sea, un objeto fractal con dimensiones entre las de una superficie y las de un volumen.
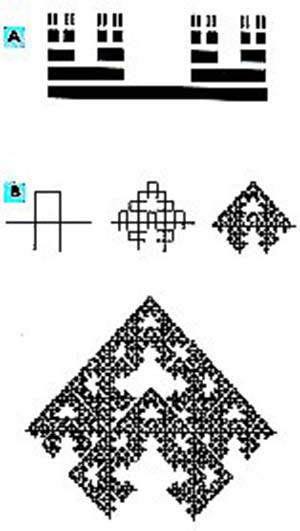
En la figura 6 se muestra, en blanco, el agregado de percolación de un fluido que se extiende hasta ocupar casi todo el reticulado. Este gráfico corresponde a una malla cuadrática de 160×160 cuadros por lado y fue obtenido por J. Feder, de la Universidad de Oslo. Un modelo de la malla de percolación es la denominada curva de Mandelbrot-Given, cuya D=.l,89 (Fig. 7B). Desde un punto de vista geométrico, un agregado de percolación ofrece mayores posibilidades de encuentro entre dos partículas, o entre un reactivo y la proteína (enzima) que cataliza su transformación. ¿Cuál es la diferencia entre un medio homogéneo 2D y uno fractal con dimensión D= 1,89?. En el primer caso es posible el movimiento browniano -así llamado porque el primero en estudiarlo fue el físico ingles E. Brown (1856-1932)-, o al azar, de las partículas, mientras que, en el segundo, dicho movimiento se da en un medio mas estructurado y, por lo tanto, con menores posibilidades de desencuentro.
A comienzos de este siglo el matemático Polya enunció un teorema según el cual la probabilidad de que un objeto (pensemos en un reactivo) que se mueve en un espacio 3D encuentre un blanco o target (pensemos en una enzima) es muy pequeña, en comparación con la que tendría el mismo objeto moviéndose en un espacio 2D. Esto lleva a pensar que esa es la función de una membrana: reducir un espacio 3D a uno 2D. Podríamos extender esta idea a que esa es, justamente, la función de este medio heterogéneo que es el citoplasma; una manera, continua o discontinua, de cambiar 3D a una dimensión menor que posibilita que enzimas y reactivos u hormonas y compuestos blanco (receptores) se encuentren en la misma dimensión geométrica y aumente, así, la velocidad de las reacciones o los efectos hormonales. El acoplamiento de mecanismos bioquímicos -como los denominados motores moleculares, conducidos por procesos químicos activos (con gasto de ATP)- con la estructura fractal del citoplasma podría implicar no sólo el aumento de la eficacia en términos catalíticos, sino también energéticos, ya que dichos motores se desplazarían con direcciones privilegiadas en 2D y no estarían sometidos al mayor azar de la 3D.
El segundo aspecto importante del modelo fractal de organización del citoplasma concierne a la probabilidad de ocupación crítica o umbral de percolación, Pc, de los sitios de la red de percolación. La importancia del umbral de percolación es que todos los puntos o sitios que pertenecen a un mismo agregado de percolación, es decir, que están conectados entre sí, alcanzan zonas muy distantes de la red. La Pc para una malla o red bidimensional cuadrática es 0,593 y para una red triangular, 0,5; por lo tanto, un cambio en la Pc puede estar reflejando una modificación en la geometría de la red. La ocupación de la red más allá del umbral de percolación implicaría que regiones alejadas del espacio quedan conectadas por pertenecer al mismo agregado de percolación (Fig. 6). Esto supondría que un comportamiento local podría extenderse a todo el agregado de percolación y, así, pasar a ser global.
Sí uno parte de la evidencia de que el citoplasma puede ser mejor descripto como un medio estructurado por la presencia de redes proteicas que como una solución acuosa donde las enzimas están disueltas, entonces, ¿cuáles son las consecuencias para la función celular, por ejemplo la velocidad de procesos acelerados por enzimas, cuando una estructura de la red de percolación cambia por procesos de montaje desmontaje de proteínas del citoesqueleto? ¿Existe alguna posibilidad de que al cambiar la estructura del citoplasma esto ocasione algún efecto de coordinación de actividades globales, como el metabolismo? Esta última pregunta adquiere especial relevancia desde el momento en que el montaje-desmontaje de elementos del citoesqueleto ocurre en las dimensiones de espacio y tiempo que podrían dar lugar a fenómenos de autoorganización y coherencia macroscópica (véase recuadro ‘La autoorganización…’).
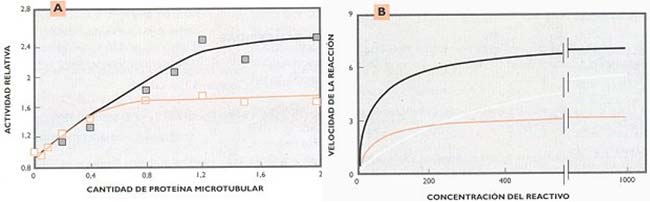
En experimentos que realizamos en colaboración con Alfredo Cáceres, del Instituto de Investigación Mercedes y Martin Ferreyra, de Córdoba, pudimos observar, tanto in vitro como en células permeabilizadas, que la velocidad de las reacciones enzimáticas aumentaba en presencia de proteína microtubular (uno de los componentes del citoesqueleto) montada o desmontada (Fig. 8). Posteriores resultados experimentales y de modelización matemática nos llevaron a postular que la distribución heterogénea de ensamblados del citoesqueleto actuaría como “fuerza conductora” de la organización dinámica de las reacciones del metabolismo celular.
Parece, entonces, muy útil explorar más a fondo la idea de que el punto de transición, que aparece al relacionar los distintos niveles de organización celular (véase recuadro ‘Niveles…’), y el umbral de percolación de los fractales proteicos del citoplasma celular estén íntimamente relacionados. En este caso sería posible que la “decisión” que toman las células de crecer o dividirse esté coordinada en el espacio y en el tiempo por procesos autoorganizados mediados por la dinámica estructural del citoplasma.




