




En los últimos años la ciencia exoplanetaria, dedicada al estudio de planetas y otros cuerpos ubicados más allá del sistema solar, viene investigando las condiciones que pudieran hacer habitable a uno de tales planetas. Por mucho tiempo se los buscó en zonas relativamente cercanas a estrellas –tanto semejantes como diferentes del Sol– en cuya superficie, por lo menos teóricamente, pudiese existir agua. Esas regiones se denominaron zonas habitables. Pero ¿qué ocurriría si eliminamos la incidencia de luz solar? ¿Sería posible que existiera un planeta habitable sin una estrella como fuente de calor? En tiempos más recientes, esta búsqueda de posibles lugares que puedan albergar vida se extendió a cuerpos fuera de las zonas habitables, por ejemplo, a planetas congelados con océanos bajo su superficie, asteroides o planetas que flotan libremente sin pertenecer a ningún sistema estelar.
Recientemente un grupo de investigadores, que publicó sus resultados en The Astrophysical Journal Letters, analizó las condiciones en las que un planeta que no recibiese luz solar podría obtener el calor necesario para albergar solventes líquidos en su superficie a partir del decaimiento de elementos radiactivos. Dichos científicos se centraron en analizar el papel que podría desempeñar tal fuente de calor en función del tamaño, de la edad y de la abundancia de elementos radiactivos de esos planetas. Incluso analizaron las posibilidades de detectar este tipo de cuerpos celestes utilizando el telescopio espacial James Webb, el sucesor del Hubble, a lanzarse en el segundo semestre de 2021.
La primera cuestión que analizaron en su publicación fue la probabilidad de tener un planeta habitable con solventes líquidos como agua, amoníaco o etano, los tres capaces de dar lugar a reacciones bioquímicas. La segunda fue establecer qué radionucleidos podría haber en un planeta de tales características, y consideraron que ellos podrían ser el uranio-238, el torio-232 y el aluminio-26, los dos primeros con largo tiempo de decaimiento y el tercero con tiempo de decaimiento mucho menor. Su análisis los llevó a concluir que planetas con una abundancia de esos radionucleidos aproximadamente mil veces mayor que la de la Tierra podrían generar el calor suficiente para que existiesen solventes líquidos en su superficie y, más aún, que tales solventes se mantengan el tiempo suficiente para que se desarrolle la vida, suponiendo que la evolución de esta requiriese un tiempo similar a la que necesitó en la Tierra.
Las condiciones así definidas implican dosis de radiación cientos de veces más altas que las padecidas por los residentes de Chernobyl durante el accidente de 1986. ¿Permitirían dichas condiciones el desarrollo de la vida? Para organismos multicelulares como los que conocemos, tanto animales como plantas, la respuesta sería no. Sin embargo, se sabe de la existencia en nuestro planeta de microorganismos, denominados extremófilos, que soportan condiciones extremas. A ese grupo pertenecen, entre otros, las bacterias Deinococcus radiodurans, que soporta dosis altas de radiación, y Desulforudis audaxviator, que obtiene sus requerimientos energéticos indirectamente de la radiación.
Si bien aún no se conocen planetas que reúnan las condiciones definidas, los investigadores sugieren que el centro de la Vía Láctea donde la colisión de estrellas de neutrones produce altos niveles de elementos pesados, como el uranio y el torio, es un buen punto para iniciar su búsqueda e incluso han identificado dos regiones del cosmos que, a pesar de estar a grandes distancias, se encuentran lo suficientemente cerca para que sean analizadas por el telescopio espacial James Webb.
Federico Gascue
[email protected]
Mayra Manente
[email protected]
Más información en LINGAM M & LOEB A, 2020, ‘On the habitable lifetime of terrestrial worlds with high radionuclide abundances’, The Astrophysical Journal Letters, 889, 20: 1-5. DOI 10.3847/ 2041-8213/ab68e5
Transferencia horizontal de genes y la colonización del medio terrestre por las plantas
Apartir del enfriamiento y la consolidación de la corteza terrestre, por cientos de millones de años (Ma) la superficie de nuestro planeta consistió de rocas estériles y desagregados finos precursores de suelos o protosuelos, moldeados por microorganismos. Las primeras plantas terrestres habrían aparecido unos 500Ma antes del presente, en la era paleozoica (541-252Ma). La proliferación de las plantas contribuyó al descenso de los niveles de dióxido de carbono (CO2) en la atmósfera y al aumento de los de oxígeno, lo cual generó un hábitat propicio para la aparición de otros grupos de seres vivos. Las plantas terrestres o embriofitas comprenden a las briofitas, como los musgos, y las plantas vasculares, que incluyen helechos y plantas con semilla (angiospermas y gimnospermas).
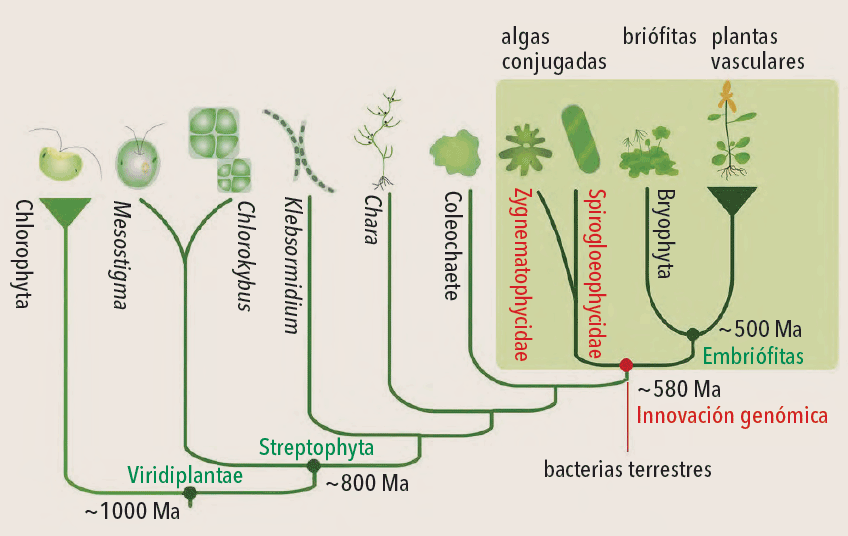
La conquista del medio terrestre tuvo lugar gracias a las adaptaciones morfológicas que permitieron a estos organismos existir en el medio gaseoso. Un estudio reciente, citado al final, sugiere que un grupo de algas verdes de estructura simple, conocidas como algas conjugadas (Conjugatophyceae y Zygnematophyceae) y todas las plantas terrestres tuvieron un ancestro común que, mucho antes de que las plantas conquistaran la tierra, vivió en un ámbito intermedio entre el acuático y terrestre, o medio subaéreo. Esto se contrapone con la hipótesis tradicional de que las embriofitas comparten un ancestro común con algas de organización más compleja, como las carofíceas (Charophyceae).
Para llegar a esta conclusión, los responsables de dicho estudio secuenciaron el genoma de dos especies de algas conjugadas (Spirogloea muscicola y Mesotaenium endlicherianum) que comparten su hábitat con musgos. Al comparar su ADN con el de diversas plantas terrestres, identificaron genes presentes y ausentes en ambos grupos. Los solo presentes en las plantas terrestres, es decir, solo aparecidos en la evolución con estas y luego diversificados con los procesos de selección natural, desempeñaron un papel fundamental en el desenvolvimiento y la expansión de ese grupo. Otros genes, adquiridos tempranamente por el ancestro común de las plantas terrestres y las algas conjugadas, resultaron indispensables para la fisiología de las plantas, pues regularon su proceso de crecimiento y sus defensas contra factores adversos, entre ellos organismos patógenos y condiciones ambientales como la desecación. Las plantas terrestres tienen también genes que condujeron al crecimiento de micorrizas, unas asociaciones simbióticas con hongos específicos del suelo que podrían haber permitido a dichas plantas obtener nutrientes de la tierra para sobrevivir. Las algas conjugadas de hábito acuático carecen de ellas.
Los autores del estudio comentado suponen que muchos de los genes esenciales para la vida fuera del agua se habrían adquirido por transferencia horizontal entre bacterias terrestres y el ancestro común de las plantas terrestres y algas conjugadas. La transferencia genética horizontal o lateral es un mecanismo frecuente entre microorganismos unicelulares, y entre estos últimos y los pluricelulares. A diferencia de la transferencia vertical, que es el pasaje de genes de ancestros a su descendencia, la horizontal es el movimiento de genes entre organismos de distintos grupos. En este segundo caso, la transferencia habría ocurrido entre bacterias terrestres y el ancestro común de las plantas terrestres y las algas conjugadas, aunque estuvieran biológicamente muy poco relacionados. Se estima que tal transferencia podría haber acontecido hace alrededor de 580Ma.
Sol Noetinger
[email protected]
Más información en CHENG S et al., 2019, ‘Genomes of subaerial Zygnematophyceae provide insights into land plant evolution’, Cell, 179, 5: 1057-1067.
HUMOR
PICHONES
Los pichones de las aves se pueden categorizar en dos grandes grupos (y todas las gamas intermedias) nidícolas y nidífugos. Los pichones nidícolas nacen pelados, ciegos, y desnudos y dependen completamente de sus padres para que los alimenten y les den calor. Al nacer, permanecen en el nido durante todo el período inicial del desarrollo. Los pichones nidífugos, en cambio, nacen con plumón, ojos abiertos, y listos para salir a pasear. Los pollitos de las gallinas, y pichones de los patos y de los teros, entre otros, son nidífugos. Los jacanas (Jacana jacana), como los de la ilustración, habitan áreas cercanas a cuerpos de agua. Caminan por la vegetación flotante, ayudados por dedos y uñas súper largas que les permiten distribuir el peso. Irene Negri





