




Nuevas metodologías e interpretaciones
El 9 de enero de 1834 durante su viaje por América del Sur como miembro científico de la expedición del Beagle, Charles R Darwin (1809-1882) descubrió en las proximidades de Puerto San Julián, en la Patagonia, ‘en el barro rojo que cubre la grava de la llanura de 90 pies’, los huesos de un mamífero extinguido sumamente enigmático. Poco después, en marzo de ese mismo año, comunicó su hallazgo por carta al naturalista inglés John S Henslow (1796-1861): ‘En Puerto San Julián encontré unos huesos muy bien preservados de un animal grande, imagino Mastodon’. El descubrimiento consistió en partes del esqueleto, incluyendo algunas vértebras y partes de los miembros, que fueron transportados a Inglaterra. Los fósiles fueron estudiados en detalle por el reconocido anatomista Richard Owen (1804-1892) del Royal College of Surgeons y finalmente resguardados en el Natural History Museum de Londres. Owen publicó la descripción de estos restos en 1838 en el libro Zoology of the Voyage of H.M.S Beagle. El animal fue nominado Macrauchenia patachonica, haciendo referencia a la longitud de las vértebras del cuello (mακρος, largo; αυχην, cuello) y a su procedencia geográfica, la región sur de América del Sur. Las extrañas características de los restos encontrados dificultaron su clasificación y Owen se refirió a ellos como ‘un gran animal mamífero, referible al orden Pachydermata, pero con afinidades de los Ruminantia, y especialmente de los Camelidae’. Esta primera apreciación, aunque un tanto confusa en el sentido de la sistemática actual, fue utilizada por Darwin para reafirmar su interpretación de que los animales actuales conservan un parentesco próximo con las formas extinguidas, relacionando guanacos y vicuñas (ambos camélidos) con la Macrauchenia extinguida. Actualmente se sabe que la relación filogenética entre ellos no es cercana, aunque de todos modos, a casi dos siglos del primer descubrimiento, Macrauchenia sigue despertando la curiosidad de los científicos del mundo entero.
¿DE QUÉ SE TRATA?
La paleontología, la anatomía comparada, las proteínas antiguas y el ADN antiguo: las controversias en la interpretación de las formas fósiles de morfologías particulares y los aportes de la biología molecular. El caso de Macrauchenia patachonica, un extraño mamífero del Pleistoceno de América del Sur.
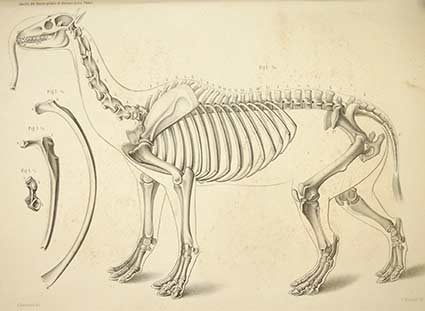
En 1856 Auguste Bravard (1803-1861), un naturalista, geógrafo y geólogo francés radicado en la Argentina, recolectó cerca de la ciudad de Salto, provincia de Buenos Aires, un cráneo casi completo y gran parte del esqueleto de Macrauchenia. Lamentablemente, Bravard murió durante el fuerte terremoto ocurrido en 1861 que destruyó casi por completo el casco de la antigua ciudad de Mendoza, antes de que pudiera completar la descripción del nuevo material. Para ello había realizado algunos excelentes dibujos que aún se siguen utilizando en la literatura científica. Bravard utilizó el nombre Opistorhinus falconeri en 1857 para referirse al fósil, aludiendo a la extraña posición de las aperturas nasales en la parte posterior del hocico, aunque este nombre quedó en desuso por tener prioridad el publicado por Owen algunos años antes.

En 1864, el reconocido paleontólogo alemán Hermann Burmeister (1807-1892), entonces director del Museo Público de Buenos Aires (actualmente MACN), describió en detalle el material descubierto por Bravard utilizando las planchas gráficas inéditas conservadas en el museo. En su artículo, Burmeister comparó reiteradamente la anatomía de Macrauchenia con los caballos y dio particular atención a la curiosa posición de las aperturas nasales, sugiriendo la posibilidad de que se vinculara a una trompa móvil. En 1889, Burmeister se ocupó nuevamente de Macrauchenia en su publicación Los caballos fósiles de la pampa argentina y realizó una primera reconstrucción, ilustrando una trompa.
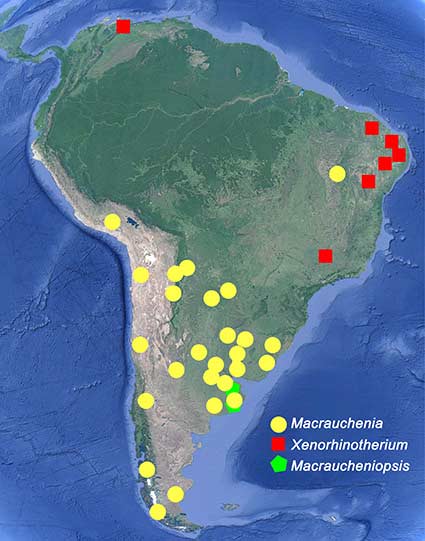
Con el paso del tiempo, otros restos de Macrauchenia fueron hallados en diferentes regiones de América del Sur. Asimismo, otros fósiles de aspecto muy similar, como Macraucheniopsis y Xenorhinotherium, fueron descubiertos, resultando en una amplia distribución a lo largo de toda América del Sur durante el Pleistoceno (de 2,6 millones de años hasta 11.700 años atrás) de estos animales (agrupados en la familia Macraucheniidae, del orden Litopterna). La historia de los macrauquénidos no se restringe al Pleistoceno, sino que también otras especies cercanamente relacionadas de los géneros Promacrauchenia, Oxyodontherium, Huayqueriana y otras fueron recuperadas de rocas más antiguas (es decir, del Mioceno tardío al Plioceno, entre 5 y 2,6 millones de años atrás). Este conjunto de especies fósiles permitió reconstruir algunos aspectos de la anatomía del grupo y proponer distintas hipótesis sobre las relaciones de parentesco con otros mamíferos, aunque resultaron controversiales en varios casos.
Tanto Macrauchenia como los otros macrauquénidos tienen un esqueleto ciertamente inusual, con las vértebras del cuello largas y robustas que recuerdan a las de un camello, los miembros locomotores con solo tres dedos como en un tapir y el cráneo robusto y alargado similar al de un caballo, aunque con las aperturas nasales ubicadas en la parte más dorsal del cráneo, vertical al paladar, similar a lo que ocurre en un delfín. La posición de las fosas nasales en la superficie dorsal (hacia arriba) del cráneo es sin duda una de las características más intrigantes de su anatomía craneana. Contrariamente, en la mayoría de los mamíferos terrestres las aperturas nasales se encuentran en el extremo del hocico y la cavidad nasal abre en ángulo recto a la cavidad nasofaríngea. En los macrauquénidos, además de la retracción de las aperturas nasales, el tamaño de las coanas es enorme. Asimismo, en la parte anterior e interna del hocico existe un saco ciego, el meatus nasi ventralis, que evidentemente representa una señal filogenética vinculada a la migración posterior de las coanas. Los avances tecnológicos, como la utilización de tomografías computadas de alta resolución, ha permitido efectuar nuevos estudios detallados sobre esta región del cráneo.
Macrauchenia: el arte y la ciencia
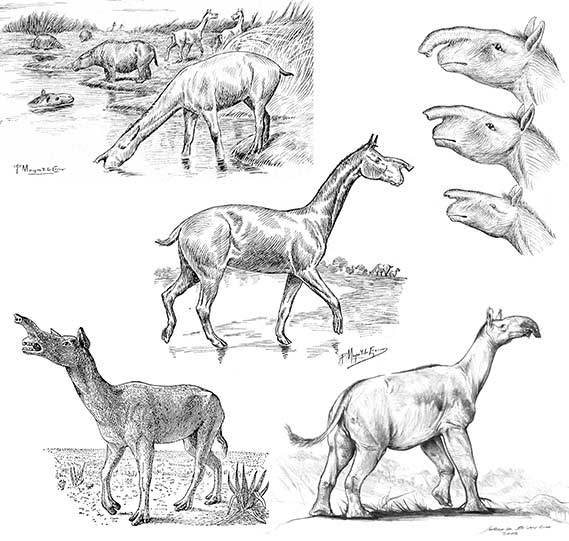
En 1925, el investigador sueco Ivar Sefve (1886-1953) realizó un trabajo sobre la estructura del cráneo de Macrauchenia, dando especial atención a la función de la respiración. Su hipótesis, por demás novedosa, implicaba la presencia de un opérculo en el techo craneano que el animal habría podido abrir o cerrar (como un delfín), implicando entonces hábitos semiacuáticos. Conforme a ello, el artista R Hellman ilustró una Macrauchenia sumergida, con sus fosas nasales cerca de la superficie del agua y otras fuera del agua.
Sin embargo, la reconstrucción de Macrauchenia con un apéndice nasal, similar a una trompa, es la hipótesis que más adeptos ha tenido. Algunas reconstrucciones muy utilizadas en la literatura fueron realizadas por el artista francés Paul Magne de la Croix (1875-1942), quien bajo la orientación de Carlos Rusconi (1898-1969), naturalista argentino y director del Museo de Ciencias Naturales y Antropológicas JC Moyano de la ciudad de Mendoza, ilustró no solo Macrauchenia sino gran parte de la fauna cenozoica argentina a principios del siglo XX. Magne de la Croix reconstruyó una trompa corta, aunque retráctil, en Macrauchenia. Por su parte Rusconi incursionó también en varios otros macrauquénidos emparentados. En sus dibujos Macrauchenia poseía una protuberancia proyectada del labio superior, siendo más reducida en los macrauquénidos más antiguos.
En el libro A History of Land Mammals in the Western Hemisphere (1913) del paleontólogo norteamericano William B Scott (1858-1947), el dibujante Robert B Horsfall ilustró una trompa larga y prensil, incluso aludiendo a que era usada para llegar a las hojas altas de la copa de los árboles. Sucesivamente, durante todo el siglo XX las reconstrucciones de Macrauchenia han tomado vida por numerosos artistas, comúnmente utilizando la estructura corporal de un camélido y una trompa en su cabeza. Reconstrucciones más recientes, como las del actual artista argentino Jorge L Blanco, ilustran una probóscide más similar a la de un tapir.
La probóscide es una extensión tubular y flexible de la musculatura tanto narial como del labio superior. En los elefantes, la trompa, conformada por una compleja musculatura intrínseca, es el análogo a una mano flexible y es manejada para asir los vegetales que consumen, transportar agua a la boca o limpiar el cuerpo. En los tapires, sin embargo, la estructura nasal es un hidrostato muscular y la función vinculada a tomar los alimentos es más limitada, si realmente existe.

Un reciente estudio que explora la función de la peculiar morfología craneana de los macrauquénidos, entre otros mamíferos, fue realizado en 2018 por los investigadores argentinos Silvana R Moyano y Norberto P Giannini a través del estudio de la anatomía comparada, utilizando como referencia el cráneo del tapir. El análisis de los caracteres seleccionados sugirió que los macrauquénidos se asemejan al alce, excepto por la presencia de grandes espacios neumáticos intracraneanos (senos paranasales), pequeños en el alce. Existe en la literatura el argumento de que los senos intracraneanos podrían favorecer la flotación de la cabeza al nadar. Sin embargo, mamíferos de grandes tallas corporales presentan casi universalmente estas cavidades, las que reducen la masa del cráneo, independientemente de la relación que tenga el animal con el medio acuático. Asimismo, resulta incierta la posibilidad de que Macrauchenia haya sido semiacuático. Cabe destacar que los fósiles de esta especie fueron hallados en distintas partes del continente, incluso en regiones que fueron áridas durante el Pleistoceno. Además, los estudios de la paleohistología de los huesos largos, que brindan información sobre la paleoecología de las especies fósiles, no son conclusivos sobre la utilización del medio acuático en Macrauchenia. En síntesis, no hay mamífero viviente con una arquitectura craneana comparable a la de los macrauquénidos. Si bien pareciera que algún tipo de apéndice nasal habría estado presente en Macrauchenia, la función es aún tema de debate.
Macrauchenia en el árbol genealógico
Dada la particular combinación de caracteres morfológicos de Macrauchenia y de otros macrauquénidos, las interpretaciones sobre su relación de parentesco con los otros mamíferos no dejaron de ser controvertidas. El paleontólogo argentino Florentino Ameghino (1854-1911) incluso pensó a fines del siglo XIX que era un perisodáctilo (Perissodactyla), esto es, un ungulado con dígitos impares como los caballos, tapires y rinocerontes. Sin embargo, incertidumbres sobre su posición filogenética han existido desde el primer hallazgo debido a la misteriosa arquitectura del esqueleto de la especie, hasta hace poco única evidencia sobre la cual basar las hipótesis de las relaciones de parentesco de las especies fósiles.
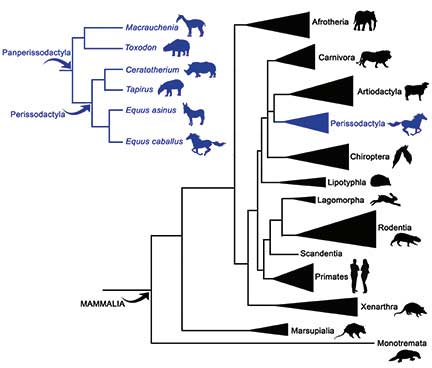
Las relaciones filogenéticas de Macrauchenia (y de otros ungulados fósiles de América del Sur, como Toxodon) recientemente fueron abarcadas, en 2015, por la biología molecular. El estudio de las proteínas fósiles, la paleoproteómica, aportó nuevas evidencias a favor de una hipótesis filogenética en particular: efectivamente, Macrauchenia y los otros miembros de su orden (Litopterna), junto con otros ungulados nativos de América del Sur, como los notoungulados, compartieron con los perisodáctilos una misma rama del árbol filogenético. Este grupo en su conjunto se reconoció con el nombre de Panperissodactyla. Los litopternas y los notoungulados se habrían diferenciado de los perisodáctilos al comienzo del Cenozoico, hace más de 60 millones de años, convirtiéndose de este modo en uno de los linajes de mamíferos nativos más antiguos de América del Sur.
LA TOMOGRAFÍA COMPUTADA
En la década de 1980, la tomografía computada de alta resolución (CT o µCT), utilizada en medicina e ingeniería, comenzó a aplicarse en estudios paleontológicos. En particular, el cráneo encierra y protege el cerebro, el cerebelo y el bulbo raquídeo, y posee diversas estructuras sensoriales como ojos, oídos, nariz y boca, por lo cual es una fuente de información formidable tanto para los estudios filogenéticos como para los paleobiológicos. Originalmente, algunos estudios intentaron acceder al interior del cráneo por medio de moldes naturales o moldes artificiales hechos en yeso o caucho, excepcionalmente representando reconstrucciones fieles.
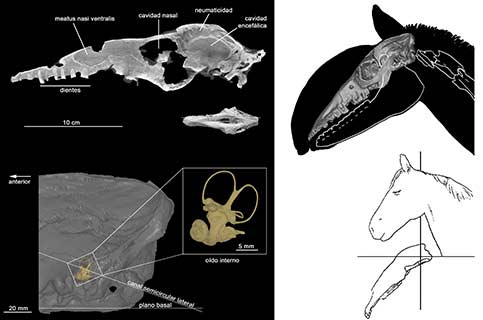
Al contrario, la tomografía computada es una técnica no destructiva y junto con la utilización de programas específicos de reconstrucción en 3D se logra acceder a las estructuras internas y generar modelos virtuales (por ejemplo, la reconstrucción de los huesos turbinales o del petroso) o el relleno de las cavidades (como por ejemplo de oído interno, cavidades neumáticas y cavidad encefálica) (ver Trotteyn MJ y Paulina-Carabajal A, 2018, ‘Paleoneurologia’, Ciencia Hoy, 27, 161: 60-64).
A pesar de que los resultados obtenidos a partir del estudio de las proteínas fósiles parecieran ser robustos, la ciencia siempre requiere la ratificación de las conclusiones a través de otras líneas de evidencias. De este modo, un segundo estudio fue abordado en 2017, esta vez a través del estudio del ADN antiguo (ADNa). En este campo, el entomólogo George O Poinar (n. 1936) en la década de 1980 fue un pionero en intentar recuperar el ADNa de insectos fosilizados en ámbar. Aunque luego se comprobó que las secuencias obtenidas oportunamente eran el producto de contaminación, sin duda captó la atención del público general en esta disciplina, inmortalizándose en la novela Parque Jurásico de Michael Crichton. En este segundo estudio sobre Macrauchenia se logró recuperar el ADN mitocondrial (ADNmt) de un espécimen encontrado en la Patagonia chilena de hace aproximadamente 11.000 años. Es frecuente que los ambientes fríos, como las regiones con permafrost, sean las más apropiadas para la preservación del material orgánico. El ADNmt es el material genético encerrado en las organelas celulares llamadas mitocondrias, se hereda solo por vía materna y resulta de utilidad para evaluar las relaciones de parentesco de las especies. Los resultados del estudio fueron coincidentes con los esperados. El análisis del ADNa dio apoyo al grupo de los Panperissodactyla, el mismo grupo obtenido a partir de la evidencia proteínica. Es interesante destacar la coincidencia pues, a diferencia del ADNmt, las proteínas del colágeno son codificadas por el núcleo celular.

Qué otros problemas se podrían abordar con el estudio de las biomoléculas? Los estudios en Macrauchenia no están terminados. Además, existen otros grupos enigmáticos en la fauna extinguida de América del Sur en los que resultaría de interés investigar o testear las hipótesis filogenéticas basadas en la morfología, como los perezosos extinguidos, recientemente estudiados con las mismas metodologías. El problema surge en que la mayoría de los grupos más controversiales se extinguieron antes del Pleistoceno y por lo tanto es muy difícil que contengan evidencia de ADNa o aun colágeno de calidad. Sin embargo, nuevas posibilidades de análisis están en progreso. Durante el desarrollo ontogenético y la formación del esmalte en los dientes, pequeñas cantidades de proteínas, llamadas proteínas del esmalte, quedan atrapadas en los minúsculos espacios interprismáticos. Estas podrían ser sujetas a análisis y la razón del interés es que se encuentran mejor protegidas de las influencias ambientales que el colágeno de los huesos. El esmalte es el tejido más duro y mineralizado del cuerpo, con el 97% de su estructura correspondiente al mineral hidroxiapatita. Por el contrario, el hueso es mucho más poroso y permite el ingreso del agua al interior de la estructura, lo que acelera el proceso de degradación del material orgánico. Los ensayos continúan, en conjunto con los perfeccionamientos en las técnicas y los programas de análisis, esperando el éxito en un futuro próximo.
LOS AVANCES EN LA BIOLOGÍA MOLECULAR Y LA PALEOPROTEÓMICA
La paleoproteómica es una disciplina molecular que estudia las proteínas antiguas. En años recientes nuevas tecnologías han permitido acceder a la información contenida en la materia orgánica preservada en los huesos fósiles. La proteína clave en la paleoproteómica es el colágeno. Este es la proteína más abundante de la piel y los huesos, por lo cual se convierte eventualmente en un recurso abundante en la paleontología. Forma fibras resistentes, por lo que tarda más tiempo en degradarse que el ADN. En condiciones excepcionales de preservación, como en el Ártico, el ADN sobrevive no más de 800.000 años. Sin embargo, el colágeno bajo circunstancias similares ha logrado recuperarse de los huesos de mamíferos del Plioceno (hace unos 3,8 millones de años). Por su parte, las proteínas pueden eventualmente preservarse en un espectro más amplio de ambientes que el ADN, si bien, al igual que este, la preservación de la molécula es mejor en condiciones ambientales frías o en suelos congelados. El colágeno es fácil de muestrear y la contaminación es más fácil de detectar que en los estudios de ADN. Con el estudio de las proteínas se puede obtener información genómica similar al ADN, aunque de alcance más limitado. Esto es porque el colágeno está formado por la actividad de algunos pocos genes, aunque la información es suficiente para establecer las relaciones de parentesco de grupos taxonómicos grandes, a diferencia del ADN, cuya resolución de trabajo es mayor: a nivel de género o especies o incluso poblacional o individual.
El procedimiento para extraer el colágeno del hueso involucra varios pasos. Primero se disuelve el material inorgánico del hueso. Luego se usa un espectrómetro de masas con un láser para fragmentar y clasificar los aminoácidos Y posteriormente se generan espectros que se comparan repetidas veces con el material de referencia de las especies vivientes.
La paleoproteómica es un campo nuevo de trabajo de utilidad en las disciplinas que estudian el pasado como la paleontología y la arqueología. En esta última ciencia se ha utilizado, por ejemplo, la técnica del colágeno para identificar los huesos de los animales que comían los hombres del pasado. La proteómica del colágeno y el esmalte tienen un potencial muy grande en la ciencia, más allá del área de la medicina de donde surgió.
En síntesis, los avances de la biología molecular aportan nuevas herramientas y metodologías de análisis con un enorme potencial para reconstruir las relaciones de parentesco de los distintos vertebrados, incluyendo las especies extinguidas, como la enigmática Macrauchenia, contrastar las hipótesis previas e intentar comprender de este modo la compleja historia evolutiva de los mamíferos fósiles de América del Sur.
LECTURAS SUGERIDAS
DELSUC F et al., 2019, ‘Ancient mitogenomes reveal the evolutionary history and biogeography of sloths’, Current Biology, 29: P2031-2042.
FORASIEPI AM, MACPHEE RDE, HERNÁNDEZ DEL PINO S, SCHMIDT GI, AMSON E & GROHÉ C, 2016, ‘Exceptional skull of Huayqueriana (Mammalia, Litopterna, Macraucheniidae) from the Late Miocene of Argentina: Anatomy, systematics, and paleobiological implications’, Bulletin of the American Museum of Natural History, 404: 1-76.
FORASIEPI AM, MARTINELLI AG Y BLANCO JL, 2007, Bestiario fósil: mamíferos del Pleistoceno de la Argentina, Albatros, Buenos Aires.
MOYANO SR y GIANNINI NP, 2017, ‘Comparative cranial ontogeny of Tapirus (Mammalia: Perissodactyla: Tapiridae)’, Journal of Anatomy, 231: 665-682.
PRESSLEE S et al., 2019, ‘Palaeoproteomics resolves sloth relationships’, Nature Ecology and Evolution, 3: 1121-1130.
WESTBURY M et al., 2017, ‘A mitogenomic timetree for Darwin’s enigmatic South American mammal Macrauchenia patachonica’, Nature Communications, 8: 15951.

Doctor (PhD) en ciencias, Universidad de Louisville, Estados Unidos.
Investigadora independiente en el IANIGLA, Conicet.

Doctor en ciencias, UFRGS. Investigador adjunto en el MACN, Conicet.

Doctor (PhD) en ciencias, Universidad de Alberta, Canadá. Curador del Departamento de Mamíferos del Museo Americano de Historia Natural, Nueva York, Estados Unidos.



