




Caenorhabditis elegans, de la naturaleza al laboratorio y del laboratorio a la naturaleza.
El premio Nobel de fisiología o medicina 2002 fue otorgado a tres científicos de la Universidad de Cambridge por sus investigaciones sobre la regulación del desarrollo de los órganos y la muerte celular programada: Sydney Brenner, John E Sulston y H Robert Horvitz. Al recibir el premio en Estocolmo, Brenner declaró, con su famoso sentido del humor, que ‘sin duda el cuarto ganador del premio Nobel este año es Caenorhabditis elegans; él merece todo el honor pero, por supuesto, no podrá compartir la recompensa monetaria’.
¿DE QUÉ SE TRATA?
Caenorhabditis elegans, un sencillo nematodo de suelo que se transformó en una herramienta fundamental para el conocimiento de aspectos básicos y generales de la biología.
¿Quién es Caenorhabditis elegans?
Caenorhabditis elegans, que desde ahora llamaremos C. elegans, es un animal microscópico que pertenece al phylum Nematoda (en castellano nematodos), uno entre más de treinta grandes grupos de animales que existen en la actualidad. Los nematodos, y otros cuatro de esos grupos, tienen el cuerpo envuelto por una cutícula más o menos rígida que limita su tamaño. Por esa razón, al crecer se despojan de ella y generan una nueva, inicialmente blanda y plegada, que luego se estira y endurece. Ese proceso se llama ecdisis o muda y los animales que lo comparten se reúnen en un gran grupo llamado Ecdysozoa (es decir animales que mudan). El grupo de ecdisozoos más numeroso en especies es el de los artrópodos (insectos, crustáceos, arácnidos, ciempiés), que incluye alrededor de un millón de especies conocidas (se considera que hay un total de cinco a diez millones aún por descubrir). Si bien se han descripto unas 25.000 especies de nematodos, se estima que su número sería millonario. Además, son numéricamente muy abundantes: puede haber millones de pequeñísimos individuos por metro cuadrado de suelo terrestre o fondo marino.

El aspecto de la mayoría de los nematodos es el de un gusano de cuerpo cilíndrico con extremos puntiagudos. Por debajo de la cutícula, compuesta principalmente por colágeno, se encuentra una capa de músculos y, en el interior, el tubo digestivo recorre todo el cuerpo. Entre los músculos y el tubo digestivo existe una cavidad llena de líquido, con relativamente pocas células, dentro de la cual se alojan los órganos reproductivos: ovarios, testículos y sus respectivos conductos. La uniformidad en la estructura de los nematodos contrasta con la diversidad de tamaños: desde alrededor de 0,4mm en algunos nematodos marinos hasta más de 8m en Placentonema gigantisima (parásito de la placenta de ballenas).
Inicialmente, los nematodos más estudiados fueron parásitos de animales y plantas. Entre los parásitos humanos, los más frecuentes son los transmitidos a través del suelo o las aguas contaminadas por sus huevos, como los grandes Ascaris lumbricoides y Ancylostoma duodenale que infectan a cientos de millones de personas; por picaduras de insectos, como las pequeñas especies que causan las filariosis linfáticas, o por comer alimentos contaminados como es el caso de Trichinella spiralis. Los parásitos de las raíces de vegetales cultivados, como las especies de Meloidogyne, causan enormes pérdidas en las cosechas. También eran conocidos los nematodos de vida libre en el suelo y en sedimentos, tanto marinos como de agua dulce. A partir de mediados del siglo XX se describieron numerosas especies de vida libre y este proceso continúa hoy con gran intensidad. En la década de 1960 una de esas especies, C. elegans, se convirtió en una nueva estrella del mundo de la ciencia y en el nematodo más popular. Para comprender por qué este nematodo de apenas 1mm de largo suscitó tanto interés, es necesario conocer a Sydney Brenner.
El ‘niño terrible’ de la biología molecular
Sydney Brenner nació en Sudáfrica en 1927 y murió –cuando aún seguía trabajando– en Singapur en 2019. Se recibió de médico en Johannesburgo y obtuvo su doctorado en la Universidad de Oxford. A poco de llegar a Inglaterra, en 1953, fue testigo de la presentación del modelo de la doble hélice del ADN por James Watson y Francis Crick, y decidió dedicar su carrera a comprender cómo funcionan los genes. En 1956 Crick lo convocó para trabajar en la Universidad de Cambridge. Usando bacterias y sus virus (los bacteriófagos), Crick y Brenner comprobaron experimentalmente sus propias hipótesis teóricas acerca de la naturaleza del código genético en 1961 y luego mostraron cómo finaliza la síntesis de proteínas codificada por el ADN. Además Brenner descubrió con François Jacob y Matthew Meselson el ARN mensajero.
Una vez comprendidos los mecanismos básicos de la biología molecular en microrganismos, Brenner se dedicó a entender el papel de los genes en formas de vida más complejas estudiando el desarrollo embrionario de los animales. El programa de investigación del Laboratorio de Biología Molecular de Cambridge de 1963, inspirado por sus ideas, consistía en elegir un organismo multicelular y tratarlo con los métodos de la genética de microbios. Este organismo debía ser fácil de cultivar en placas de agar con bacterias como alimento, tener un ciclo de vida breve, ser pequeño para poder trabajar con muchos individuos a la vez, tener pocas células cuya genealogía se conociera, y que en él fuera posible detectar, manipular y estudiar mutaciones. El elegido fue el nematodo C. elegans que tiene, además, otra ventaja: un número constante de células somáticas (es decir, de todas las células del cuerpo excepto las células germinales destinadas a formar los óvulos y espermatozoides).
Brenner divirtió, pero también ofendió a veces a quienes lo escuchaban o leían, con sus ironías y su rechazo a dogmas y convencionalismos. Así se hicieron famosas sus columnas en la revista Current Biology que firmaba como ‘Tío Sid’.
C. elegans antes de conocer a Sidney Brenner
Caenorhabditis elegans fue descubierto en una muestra de humus en Argelia en 1897 y bautizado como Rhabditis elegans por Émile Maupás (1842-1916), un zoólogo francés que publicó sus observaciones en 1900 como parte de un extenso trabajo sobre ‘los modos y las formas de reproducción de los nematodos’. Ocurre que los nematodos varían en la forma de reproducción: en algunas especies hay machos y hembras (en general de mayor tamaño que los machos), pero también existen especies hermafroditas (ambos sexos en el mismo individuo) e incluso que presentan partenogénesis (desarrollo de embriones a partir de óvulos sin fecundar).
En la década de 1940, Victor Nigon (1920-2015), también francés, profundizó las investigaciones sobre la reproducción y el proceso de determinación del sexo en los pequeños nematodos de vida libre conocidos como rabdítidos, incluido R. elegans. En colaboración con Ellsworth C Dougherty (1921-1965), de la Universidad de California en Berkeley, Nigon describió una especie muy similar a R. elegans, a la cual llamaron Rhabditis briggsae. Esta junto con otras especies fueron incluidas en el género Caenorhabditis en 1952, y adquirieron así sus nombres actuales: C. elegans y C. briggsae. Dougherty y Nigon siguieron colaborando y describieron la primera mutación morfológica de C. briggsae. Durante la década siguiente, Dougherty logró obtener cultivos axénicos (sin ningún otro organismo vivo) de esos nematodos en placas de agar, y mantuvo poblaciones puras en el laboratorio.
Inicialmente, Brenner eligió a C. briggsae como el organismo a utilizar en sus experimentos, pero luego de visitar a Dougherty en California, eligió C. elegans. Posiblemente eso se debió a diferencias en el crecimiento y comportamiento entre ambos que hacían a C. elegans más fácil de cultivar y manipular. En octubre de 1963 un cultivo axénico de una cepa originalmente recolectada en Bristol (Reino Unido) se envió desde los Estados Unidos al laboratorio de Brenner. Esa cepa fue designada N2. Esos ejemplares fueron los antepasados del ganador del premio Nobel.
No todo empezó con C. elegans
El punto de partida del uso de C. elegans en la investigación genética puede ubicarse simbólicamente en una breve nota de la revista Nature de 1948, en la cual Dougherty y Hermione G Calhoun expusieron sus ventajas. Sin embargo, los trabajos pioneros sobre el desarrollo en nematodos fueron realizados en los grandes ascáridos parásitos de animales por el belga Édouard van Beneden (1846-1910) en la década de 1880 y el alemán Theodor Boveri (1862-1915), junto a su estudiante graduada y luego esposa Marcella O’Grady (1863-1950). Ellos describieron, a partir de 1888, la fertilización, división celular y desarrollo embrionario temprano del gran Parascaris equorum que vive en el intestino de los caballos y mide 50cm (entonces se llamaba Ascaris megalocephala).
Sus descubrimientos condujeron, nada más y nada menos, a la teoría cromosómica de la herencia. O’Grady no figuró como coautora de los trabajos, pero su papel fue clave debido a su experiencia adquirida en el laboratorio de Edmund Wilson en los Estados Unidos. Ese zoólogo construyó una especie de árbol genealógico celular de un gusano marino a partir del cigoto (la primera, producto de la fertilización del óvulo por el espermatozoide), en el cual nombró a cada descendiente mediante una práctica terminología. Los Boveri también diseñaron una terminología para cada generación de células de P. equorum, que fue publicada en 1899 y aún se usa, con algunas variaciones, incluso en C. elegans.
El ciclo de vida de C. elegans
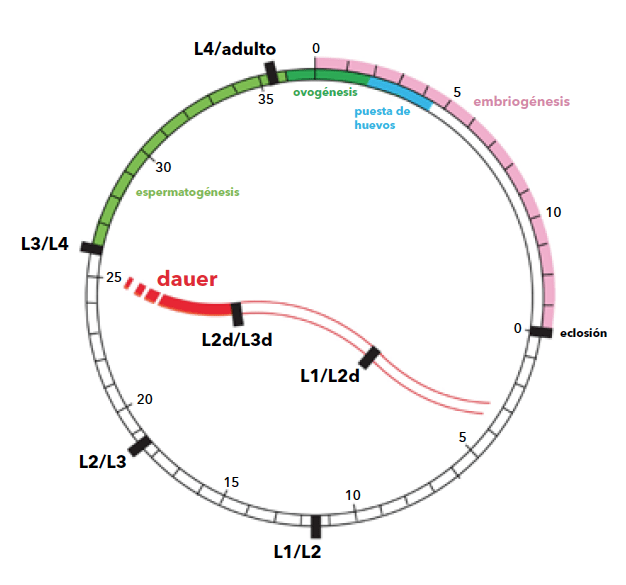
Los biólogos han acuñado el término ciclo de vida para describir los eventos que se suceden entre una generación y la siguiente: nacimiento, desarrollo, madurez y reproducción.
Los individuos adultos de C. elegans son hermafroditas que se fecundan a sí mismos y machos ocasionales que pueden fecundar a los primeros. En los hermafroditas las células germinales originan hasta 350 espermatozoides en el último estadio inmaduro y solo óvulos en los adultos. El cambio es irreversible y es el número de espermatozoides el que limita el número de huevos. En el laboratorio, aproximadamente uno de cada mil individuos se desarrolla espontáneamente como macho, pero esa proporción puede aumentar con el incremento de la temperatura. Los machos tienen un número de espermatozoides hasta tres veces mayor que los hermafroditas y por eso, cuando un macho insemina un hermafrodita, la descendencia aumenta hasta más de mil huevos. La mayoría de los hijos de un hermafrodita autofecundado son también hermafroditas y genéticamente idénticos: son clones. El cruzamiento entre un hermafrodita y un macho permite entonces un cierto grado de variabilidad genética.
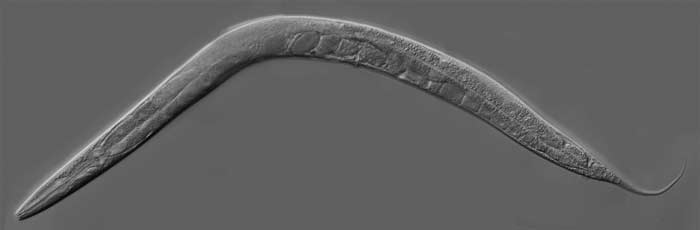
De la eclosión de los huevos surgen individuos minúsculos, similares a sus progenitores, llamados larvas o juveniles. Es el primero de cuatro estadios necesarios para llegar a la etapa adulta. En promedio, el tiempo de este proceso es de apenas dos o tres días, y la vida del adulto en el laboratorio, a una temperatura constante de 20°C, es de dos a tres semanas. Como muchos otros nematodos relacionados, el segundo estadio larval puede entrar en una vía alternativa de desarrollo en el caso de sobrepoblación o falta de alimento. Esta vía consiste en un tercer estadio larval modificado, llamado dauer (duración, en alemán), que puede sobrevivir sin alimentos varios meses y que resiste la deshidratación y otros estreses ambientales. Esta especie de letargo se conoce como diapausa y finaliza cuando las condiciones ambientales son favorables; entonces el ciclo se reinicia mudando a un cuarto estadio normal. En otras palabras, C. elegans tiene mecanismos que le permiten alargar su vida.
Linajes celulares de C. elegans y la muerte celular programada
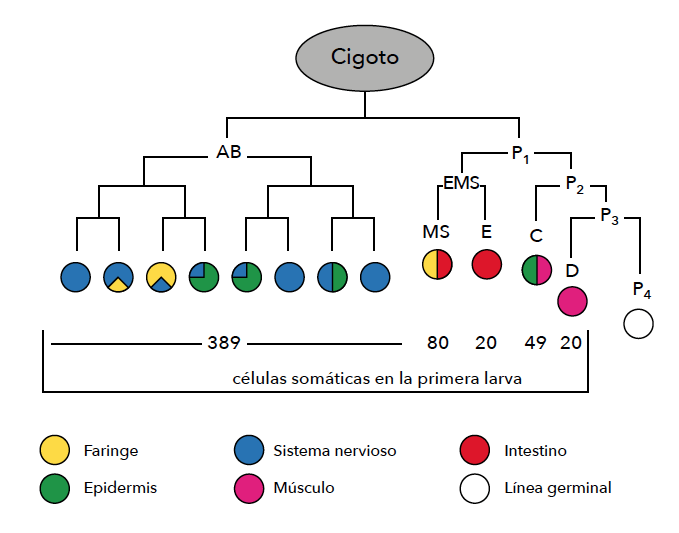
Los linajes celulares de C. elegans desde el cigoto hasta la larva que eclosiona del huevo fueron revelados en detalle en 1983 por el biólogo británico John E Sulston (1942-2018), que compartió el Nobel con Brenner. En ese proceso se generan 671 células; en un hermafrodita 113 de ellas (en los machos 111) mueren de forma genéticamente programada, un proceso que se llama apoptosis. Las restantes se diferencian de forma definitiva o lo harán durante el posterior desarrollo larval.

La primera división celular es asimétrica, y origina una célula fundadora grande (AB) y una célula madre más chica (P1). AB y P1 además difieren en sus destinos celulares: AB origina exclusivamente células somáticas y P1, tanto células somáticas como germinales.
El número de células se duplica durante las siguientes divisiones celulares, pero interesa analizar la descendencia de P1, que da origen a una nueva célula fundadora somática y una nueva célula madre, P2. Lo mismo ocurre con P2, que se divide y da una tercera célula fundadora somática, y la célula madre P3. Finalmente, P3 se divide en una célula fundadora somática y en P4, que permanece en reposo hasta el último estadio larval y es la madre de todas las células reproductivas del adulto.

Con anterioridad, en 1977, Sulston y Horvitz habían trazado la genealogía de las 959 células somáticas del hermafrodita adulto y las 1031 del macho, que se originan de la división de las 558 o 560 células del primer estadio larval menos 131 o 148 células muertas por apoptosis, respectivamente.
El primer genoma secuenciado de un organismo multicelular fue el de C. elegans
A lo largo de todo el siglo XX se llevó a cabo uno de los programas de investigación más importantes de la historia de la biología que permitió entender la naturaleza del gen, gracias al descubrimiento de la estructura del ADN en 1950.
La tapa de la revista Science del 11 de diciembre de 1998 mostró una foto de C. elegans y en su interior incluyó un artículo firmado por una asociación de unos cuatrocientos investigadores en el que se describe la secuencia de su genoma, es decir, el orden en que se encuentran los 97 millones de pares de bases del ADN que forman sus más de 19.000 genes. Fue el primer genoma conocido de un organismo multicelular, luego de los correspondientes a virus, varias bacterias y una levadura. Este genoma reveló información acerca de las bases generales de la construcción animal y las más específicas de la construcción de un nematodo. Más del 40% de las proteínas coinciden con las de otros organismos, incluyendo a los seres humanos, y la distribución de los genes en los seis pares de cromosomas fue una evidencia de la organización regional de estos.
Cinco años más tarde se publicó la secuencia del genoma de su primo hermano C. briggsae, con quien comparte el hábitat y el modo de reproducción hermafrodita, lo cual permitió refinar el conocimiento de C. elegans y es una fuente prometedora de comparaciones evolutivas.
Es hora de dejar el laboratorio y volver a visitar el campo
Los miles de millones de C. elegans que pasaron sus vidas en laboratorios del mundo provienen de una sola cepa: la N2 de Bristol, usada primero por Brenner y sus amigos, quienes las alimentaron y cuidaron. Pero ¿cómo sería la vida de esta especie en la naturaleza, enfrentando con su aparente uniformidad genética variaciones en la disponibilidad de alimento y temperatura y la presencia de depredadores y parásitos? Esa pregunta despertó la curiosidad de Marie-Anne Félix, del Instituto Jacques Monod de París.
Aunque se lo conoce como nematodo del suelo, quizá debido al sitio de su primer hallazgo, la mayoría de las veces C. elegans es encontrado en vegetales en descomposición en todos los continentes menos la Antártida. La zoóloga parisina encontró poblaciones abundantes de C. elegans y C. briggsae en antiguas plantaciones de árboles frutales de los tranquilos alrededores de su ciudad. Cientos de gusanos de todos los estadios del ciclo de vida, incluyendo los escasos machos, vivían en frutas, flores y tallos herbáceos en putrefacción. Ella y sus colegas investigaron en ambientes similares de toda Francia en las cuatro estaciones y pudieron así informar sobre aspectos de su alimentación, densidad, reproducción, comportamiento y sus relaciones con otros integrantes de su comunidad, como depredadores y patógenos, a distintas escalas espaciales y temporales.
Un aspecto fundamental de C. elegans en la naturaleza tiene que ver con la longeva larva dauer que se encuentra en material vegetal podrido, pero también sobre muchos invertebrados, incluso dentro de caracoles, cuando la densidad poblacional es extrema. Esos animales y sus predadores, como las aves, les permiten dispersarse lejos de su sitio de origen. Tal transporte, además de una mayor presencia de machos en la naturaleza, podría explicar la insospechada variación entre poblaciones de la especie, como rasgos de comportamiento e, incluso, diferencias en los linajes celulares. Esas variaciones indican mayor diversidad genética que la presente en el laboratorio.
Los estudios de laboratorio y del campo se ponen en contacto
Hay una paradoja interesante en la ciencia de la vida: existen dos áreas de investigación que, hasta hace relativamente poco tiempo, se diferenciaban de forma aparentemente irreconciliable en su método. La biología se ocupa tanto de la unidad de los procesos de la vida (que se estudia principalmente en el laboratorio) y la diversidad de los seres vivos (que se estudia sobre todo en el ambiente). La base molecular de ambos procesos es la misma: de una manera general, el ADN puede conservarse y variar. La cronología de las investigaciones que incluyen a C. elegans abarca ambos métodos y una posible conciliación de ellos. A comienzos del siglo XX, esto se advirtió en una de las varias especies de nematodos del suelo que se describieron y en las que se estudiaron los mecanismos reproductivos (diversidad). A partir de 1960, en un espectacular desarrollo de los trabajos de genética, biología celular, molecular y del desarrollo, incluyendo la muerte celular programada utilizando una única cepa de C. elegans como modelo experimental (unidad). Y por último, en un regreso al ambiente combinando los muestreos de campo con los métodos más avanzados de la biología molecular y del desarrollo (unidad más diversidad).
Caenorhabditis elegans en la Argentina
El género Caenorhabditis no ha sido encontrado en ambientes naturales de la Argentina, pero muchos laboratorios locales utilizan la cepa N2 de C. elegans en investigaciones sobre temas básicos de la biología como estrés, envejecimiento, neurobiología y ritmos biológicos, así como en farmacología, toxicología y efecto de pesticidas. Prueba de ello son los importantes trabajos publicados en revistas científicas de primer nivel por investigadores de las universidades de Buenos Aires, Córdoba, Rosario, Quilmes, San Martín y del Sur, y de los institutos del Conicet asociados a ellas.
El autor agradece a Marie-Anne Félix por el generoso envío de sus fotos para ilustrar este artículo.
Lecturas sugeridas
DEUTSCH J, 2009, El gusano que usaba el caracol como taxi, FCE, Buenos Aires.
FIDALGO M y MUÑOZ MJ, 2006, ‘Caenorhabditis elegans: los mecanismos genéticos de la longevidad’, Investigación y Ciencia, 363: 75.
JURADO P, 2013, ‘Un gusano revela claves de la memoria’, Mente y Cerebro, 60.
PORTMAN DS, 2019, ‘La mente de dos gusanos’, Investigación y Ciencia, 517: 13.
WORM BOOK, ‘An online review of C. elegans biology’. Accesible en wormbook.org

Doctor en ciencias biológicas, UBA.
Investigador principal jubilado, Conicet.
Profesor titular jubilado, UNMDP.



